

Optimization of Culture Conditionsand Analysis of Fermentation Process for Plant Vaccine-producing Bacterium, FJAT-1458
-
摘要: 对番茄青枯病植物疫苗菌株FJAT-1458进行培养条件优化和发酵过程状态参数研究。采用摇瓶培养和单因素试验研究菌株FJAT-1458的培养条件,确定最适培养基为SPA(蛋白胨5 g·L-1,蔗糖20 g·L-1,KH2PO4 0.5 g·L-1,MgSO4 0.25 g·L-1),最适培养温度为30℃,250 mL三角瓶最适装液量为35 mL。用50 L发酵罐发酵FJAT-1458,发酵过程中菌体数量呈现先升后降的变化趋势,FJAT-1458的生长可分为4个时期:0~6 h为发酵适应期、6~36 h为对数生长期、36~60 h为稳定生长期,72 h后为消亡期。发酵过程中,菌体的致病性指标-弱化指数变化小,介于0.832~0.855,方差分析表明其差异不显著,P>0.05;发酵液pH值从7.23上升至8.78;菌体细胞为短杆状,大小为(1.0~1.5)μm×(0.5~0.8)μm,随着发酵进程,细胞颜色变深,发酵后期细胞开始变形、裂解。Abstract: Conditions to culture FJAT-1458, a bacterium that produces plant vaccine against the tomato bacterial wilt caused by Ralstonia solanacearum, were optimized by asingle factor experiment. They were determined to be the use of an SPA medium containing peptone 5 g·L-1, sugar 20 g·L-1, KH2PO40.5 g·L-1, and MgSO40.25 g·L-1 at 30℃ with 35 mL of broth in a 250 mL shaking flask. Subsequently, in a 50 L fermentation tank, the fermentation was carried out and the process monitored. During the fermentation, the cell count on FJAT-1458 increased initially and followed by a gradual decline with a 4-stage growth period that included the adaptive (0-6 h), logarithmic (6-36 h), stationary (36-60 h), and decline phases (after 72 h). In addition, it was observed that(a) the pathogenicity indicator, attenuation index, of FJAT-1458 ranged from 0.832 to 0.855 showing no significant difference (P>0.05) throughout the entire process; (b) the pH of the fermentation broth increased from 7.23 to 8.78; (c) morphologically, the bacteria appeared to be(1.0-1.5)μm×(0.5-0.8)μm rods with a darkening color as the process proceeded; (d) the cellular secretion from the bacteria increased with time; and, (e) some of the bacterial cells deformed and lysed near the end of fermentation.
-
0. 引言
【研究意义】水稻是重要的粮食作物之一,我国一半以上的人口以稻米为主粮,水稻育种生产与国家粮食安全密切相关[1]。恢复系在水稻杂交育种中发挥着重要作用,恢复力强、配合力高、丰产稳产性好、抗性强、品质优、株叶形态好的恢复系在生产上有重要的应用价值[2]。杂交后代的产量和米质与恢复系直接相关,杂交稻要获得高产,其亲本要有优良的农艺性状[3]。恢复系的优良性状在选育超高产杂交水稻方面起关键作用。因此,研究分析恢复系的相关性状对其育种应用及解析杂交后代的遗传方面均有重要意义。【前人研究进展】水稻粒型是决定千粒重进而影响水稻产量的重要性状[4],同时,粒形与稻米的外观品质、加工品质和蒸煮品质也紧密相关[5-8]。研究表明,粒形受加性效应影响较大,且恢复系影响大于不育系,其中粒长的恢复系亲本方差贡献率较大[9]。已克隆的水稻粒形相关基因有GL7、GS3、GW8、SGL、GIF1等,其中水稻GL7编码的蛋白与拟南芥中LONGIFOLIA蛋白同源,调控细胞的纵向伸长[10];GS3编码包含4个结构域的跨膜蛋白,负向调控籽粒的大小和重量[11];GW8编码包含SBP结构域的转录因子,正调控细胞增殖,同时控制谷粒大小、粒型和稻米品质[12];SGL编码的蛋白是驱动蛋白13亚家族的成员,正调节细胞纵向伸长,调控籽粒长度[13];GIF1编码的蛋白是一种细胞壁转化酵素,在水稻籽粒发育时,控制着蔗糖运输卸载和灌浆,影响籽粒的充实度[14]。另外,稻米品质受环境的影响,但主要受遗传控制。杂交后代的整精米率与恢复系的整精米率呈显著正相关 [15];并且恢复系对F1代的垩白度和直链淀粉含量的影响也大于不育系[16,17]。水稻中控制米质的相关基因有Wx、Chalk5、ALK、OsSSI、OsBEIIb等,其中Wx编码颗粒淀粉合成酶,是控制直链淀粉合成的主效基因;Wx的第一内含子剪切效率与第一内含子+1的碱基直接相关,当其第一内含子+1位碱基为G时(即为等位基因Wxa),能够正常剪接,产生较多的Wx成熟mRNA进而翻译成Wx蛋白并使稻米中直链淀粉含量较高;而当第一内含子+1位碱基为T时(即为等位基因Wxb),无法正常剪接,产生保留第一内含子的mRNA前体,Wx蛋白翻译受阻,稻米中直链淀粉含量较低[18]。Chalk5编码液泡膜质子转运焦磷酸酶,具无机焦磷酸水解活性和质子转运活性,影响水稻籽粒垩白的形成和精米率等品质性状[19];ALK 基因编码可溶性淀粉合酶Ⅱ,主要控制稻米的糊化温度[20];OsSSI编码淀粉合成酶Ⅰ,影响水稻籽粒中支链淀粉的结构,从而影响稻米的糊化温度[21];OsBEIIb编码淀粉分支酶IIb,其表达水平的改变影响淀粉的结构,从而影响垩白等[22]。【本研究切入点】恢复系福恢676是在产量和品质相结合上取得突破性进展的强优势恢复系,由籼粳交中间材料 8L124与优良恢复系明恢 63杂交,再与优良恢复系蜀恢 527复交后育成[23]。现已利用福恢676配组育成了18个通过省级或国家品种审定的杂交稻品种,且福恢676配制的杂交水稻品种具有产量高、米质优、抗性好等特点。但目前关于具体分析恢复系福恢676粒形和米质及相关基因表达等方面的研究还尚未有报道。【拟解决的关键问题】本研究以恢复系福恢676、亲本明恢63和蜀恢527以及其他两个生产上常用的恢复系明恢86和福恢673为材料,分析比较其粒形及米质,确定Wx基因第一内含子+1 位碱基类型,并获得控制粒形和米质相关基因的表达模式,为恢复系福恢676的应用推广提供理论参考。
1. 材料与方法
1.1 试验材料
福恢676、明恢63、蜀恢527、明恢86、福恢673种子由福建省农业科学院农业农村部华南杂交水稻种质创新与分子育种重点实验室保存。2021年分别种植于福州、泉州和三明水稻试验基地,收获成熟种子并自然晒干,备用。
rTaq DNA聚合酶、Acc I限制性内切酶购自TaKaRa公司;总RNA提取试剂Trizol购自全式金生物技术有限公司;cDNA链反转试剂盒(东洋纺反转录试剂盒)购自东洋纺(上海)生物科技有限公司;荧光定量试剂Rox Reference DyeII购自罗氏(Roche)公司。
1.2 试验方法
1.2.1 稻谷长、宽、厚的测量
随机取适量各恢复系的种子,采用游标卡尺测量稻谷的长、宽、厚,每个材料3次重复(50粒·次−1),并测定千粒重。
1.2.2 PCR扩增及酶切分析
采用CTAB(十六烷基三甲基溴化铵)法提取水稻植株基因组DNA。以DNA为模板进行PCR扩增,所用上游引物Wx-F:5'-CTTTGTCTATCTCAAGACAC-3',下游引物Wx-R:5'-TTTCCAGCCCAACACCTTAC-3'[24];PCR反应体系:rTaq Buffer 1 µL,dNTP (2.5 mmol · L−1)0.8 µL,上游引物和下游引物(10 μmol · L−1)各 0.4 µL,DNA模板0.5 µL,rTaq 0.1 µL,ddH2O补足至10 µL。PCR扩增程序:94 ℃预变性5 min后,94 ℃ 30 s,62 ℃ 30 s,72 ℃ 30 s,30个循环,72 ℃延伸7 min。以广恢128为对照,反应完成后进行琼脂糖凝胶电泳检测,并送测序,进行序列分析。取上述PCR产物,进一步采用Acc I进行酶切分析,反应体系为:PCR产物3 µL,10× M buffer 1 µL,Acc I 0.5 µL,ddH2O补足至10 µL,37 ℃ 酶切4 h;再取5 μL 酶切产物进行琼脂糖凝胶电泳检测。
1.2.3 米质测定
收获的成熟种子,室温放置3个月。每个材料称取200 g,采用《中华人民共和国农业农村部标准米质测定方法》(NY/T 2334—2013),对样品的糙米率、精米率、整精米率、垩白率、垩白度、透明度、碱消值、直链淀粉含量、胶稠度进行测定,每个材料3次重复。
1.2.4 荧光定量PCR
在水稻扬花期,剪去已经开花和还未开花的颖花,留下当天开花的颖花,并在灌浆4 d、7 d和11 d时取样,去除颖壳,液氮速冻,保存于−80 ℃备用。采用Trizol法提取水稻种子总RNA;按照东洋纺反转录试剂盒的操作说明书,反转录合成cDNA一链,并稀释20倍。进行SYBR Green I荧光定量PCR(qRT-PCR),以水稻肌动蛋白基因Actin150为内参基因,反应体系:Rox Reference DyeⅡ 10 µL,上下游引物(10 µmol· L−1)各 0.6 µL,cDNA 2 µL,ddH2O补足至20 µL。采用Thermo实时荧光定量PCR仪(QuantStudioTM 7 Flex System),每个反应设4次重复,反应程序:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 60 s, 40个循环;反应结束后进行熔解曲线分析,熔解曲线根据标准程序进行。所用引物详见表1。
表 1 qRT-PCR引物Table 1. Primers for qRT-PCR基因名称Gene names 引物序列(5′-3′)Primer sequence(5′-3′) GL7 F: AAGGACTTCAGGGCTCTCAGGATAC R: GCTGGAAGTGTCTGGAACTGGTGTT GS3 F: GGGTGAAATAAATTCAATCGAAGGG R: GCACAAACAGCGAAACTTCTTCAAG GW8 F: AGGAGTTTGATGAGGCCAAG R: GCGTGTAGTATGGGCTCTCC SGL F: CTCTTCTATGGAACCTGACAG R: CTGAGAAGCTGAAGCAGATG GIF1 F: TTGGTAGTAGGGTCGCTTGGGC R: CCAGTCCGGAGCTCAGAGTCGA Chalk5 F: GCCGTCACCTTCCTCTCCCTCC R: CGTAAGCATTGCTCGTGAAGTACTCG ALK F: CCTATTCCTGCGGTAGAAGA R: CCGAATCGTCATCCTGGT OsSSI F: GTGAGCAGGAGTCTGAGAT R: TGACCACGAAGAGCAAGA OsBEIIb F: CGGTTTCAGCAGGTTCAGA R: CTCCAGATGACTCAATCTCAACTT Actin150 F: AGTGTCTGGATTGGAGGAT R: TCTTGGCTTAGCATTCTTG 1.3 数据分析
采用单因素方差分析或t检验对获得的数据进行统计分析,P≤0.05为差异显著;采用 SPSS 软件及Microsoft Excel 2019进行统计分析及作图。
2. 结果与分析
2.1 稻谷粒形及千粒重分析
对福恢676、亲本明恢63和蜀恢527,以及籼型恢复系明恢86和福恢673的谷粒粒形进行测量和分析。结果(图1、表2)显示,福恢676的谷粒长大多为10~11 mm,种植于3个地区的福恢676的谷粒长均介于亲本明恢63与蜀恢527之间,并且福恢676的谷粒长均大于其他两个恢复系明恢86和福恢673。福恢676的谷粒宽和谷粒厚分别为2.70 mm和2.00 mm左右,但种植于不同地区福恢676的谷粒宽和谷粒厚与其他恢复系相比没有明显的统一趋势。种植于福州和泉州的福恢676的长宽比为3.90和3.92,种植于三明的为3.72,种植于3个地区的福恢676的长宽比均明显小于蜀恢527但明显大于明恢63、明恢86和福恢673。福恢676的千粒重为27~31 g,种植于福州和三明的福恢676的千粒重均大于亲本明恢63、明恢86和福恢673,但均小于亲本蜀恢527。
![]() 图 1 不同恢复系稻谷粒形(福州)Figure 1. Shapes of rice grains of different restorer lines (Fuzhou)表 2 稻谷粒形和千粒重分析Table 2. Grain shape and 1000-grain weight of rice restorers
图 1 不同恢复系稻谷粒形(福州)Figure 1. Shapes of rice grains of different restorer lines (Fuzhou)表 2 稻谷粒形和千粒重分析Table 2. Grain shape and 1000-grain weight of rice restorers种植地区Growing areas 恢复系Restorer lines 谷粒长Grain length/mm 谷粒宽Grain width/mm 长宽比Length-width ratio 谷粒厚Grain thickness/mm 千粒重1000-grain weight/g 福州 Fuzhou 福恢676 Fuhui676 10.57±0.33 2.72±0.10 3.90±0.18 2.09±0.07 27.66±0.47 明恢63 Minghui63 9.98±0.27* 2.72±0.09 3.67±0.15* 1.99±0.05* 26.82±0.17* 蜀恢527 Shuhui527 11.28±0.34* 2.73±0.11 4.14±0.19* 2.08±0.06 29.40±0.13* 明恢86 Minghui86 9.91±0.24* 2.75±0.09 3.60±0.11* 1.94±0.10* 26.60±0.20* 福恢673 Fuhui673 10.15±0.36* 2.74±0.08 3.70±0.13* 2.00±0.07* 26.42±0.26* 泉州 Quanzhou 福恢676 Fuhui676 10.80±0.40 2.76±0.07 3.92±0.15 2.04±0.09 30.31±0.02 明恢63 Minghui63 10.51±0.25* 2.79±0.07 3.77±0.10* 2.00±0.07 30.17±0.13 蜀恢527 Shuhui527 11.54±0.39* 2.79±0.09 4.14±0.15* 2.00±0.09 31.82±0.01* 明恢86 Minghui86 9.98±0.28* 2.78±0.10 3.60±0.14* 2.06±0.07 29.44±0.38* 福恢673 Fuhui673 10.31±0.42* 2.74±0.11 3.77±0.22* 2.08±0.06 30.35±0.29 三明 Sanming 福恢676 Fuhui676 10.36±0.46 2.79±0.09 3.72±0.21 2.07±0.05 29.13±0.17 明恢63 Minghui63 9.84±0.33* 2.80±0.09 3.52±0.18* 1.99±0.08* 28.52±0.14* 蜀恢527 Shuhui527 10.85±0.32* 2.79±0.09 3.90±0.19* 2.01±0.08* 30.87±0.20* 明恢86 Minghui86 9.44±0.29* 2.85±0.12 3.32±0.19* 1.94±0.08* 27.88±0.48* 福恢673 Fuhui673 9.99±0.40* 2.86±0.10* 3.49±0.16* 1.97±0.11* 26.89±0.38* *表示差异达显著水平(P≤ 0.05),没有标注*表示没有显著差异。下表同。*indicate significant differences at P≤0.05. Same for below. 2.2 Wx第一内含子+1位碱基类型分析
以福恢676、明恢63、蜀恢527、明恢86、福恢673植株的基因组DNA为模板,用引物Wx-F/Wx-R进行PCR扩增,电泳检测显示样品中均扩增出大小为253 bp的目的条带(图2A)。进一步采用限制性内切酶Acc I 对 PCR 产物进行酶切,电泳结果显示对照广恢128的Wx片段被切成约126 bp的片段,即其Wx基因第一内含子+1位碱基为G;而5个恢复系中扩增出的Wx片段均没有被切动,仍为 253 bp(图2B),即Wx基因第一内含子+1位碱基为T而不是G[25]。进一步的测序分析也表明,5个恢复系中Wx基因第一内含子+1位碱基均为T(图3),即Wx基因型为Wxb。
![]() 图 2 Wx基因片段的PCR扩增及限制性酶切分析A为PCR扩增,B为限制性酶切分析;M为Marker 2000,1为对照广恢128,2为明恢86,3为明恢63,4为福恢676,5为福恢673,6为蜀恢527。Figure 2. PCR amplification and restriction enzyme analysis of Wx gene fragmentA: PCR amplification; B: restriction enzyme analysis; M: Marker 2000 bp; 1: Guanghui128 as control; 2: Minghui86; 3: Minghui63; 4: Fuhui676; 5: Fuhui673; 6: Shuhui527.
图 2 Wx基因片段的PCR扩增及限制性酶切分析A为PCR扩增,B为限制性酶切分析;M为Marker 2000,1为对照广恢128,2为明恢86,3为明恢63,4为福恢676,5为福恢673,6为蜀恢527。Figure 2. PCR amplification and restriction enzyme analysis of Wx gene fragmentA: PCR amplification; B: restriction enzyme analysis; M: Marker 2000 bp; 1: Guanghui128 as control; 2: Minghui86; 3: Minghui63; 4: Fuhui676; 5: Fuhui673; 6: Shuhui527.![]() 图 3 Wx基因片段的序列比对分析方框内为碱基差异位置。Figure 3. Sequence alignment on Wx gene fragmentPosition of base difference is showed in box.
图 3 Wx基因片段的序列比对分析方框内为碱基差异位置。Figure 3. Sequence alignment on Wx gene fragmentPosition of base difference is showed in box.2.3 米质分析
表3中的米质分析结果显示,福恢676及其他4个恢复系的糙米率都大于81%,均达到一级米标准;福恢676的精米率为69%~74%。福恢676的整精米率只有种植于泉州的高于55%,达到一级米标准,但种植于3个地区的福恢676的整精米率均高于亲本明恢63与蜀恢527以及明恢86、福恢673,除了种植于福州的明恢86,其他的差异均达到显著水平。长、宽及长宽比方面,福恢676与其他恢复系相比较的情况与稻谷粒形分析情况类似。种植于泉州和三明的福恢676的垩白度均值分别为3.60%和3.67%,达到三级米标准;福恢676的垩白率和垩白度均高于亲本明恢63与蜀恢527,但均低于恢复系明恢86和福恢673,且绝大部分差异都达到显著水平。种植于泉州和三明的福恢676的透明度为2,达到二级米标准。福恢676的碱消值为4.3~4.6,而直链淀粉含量均处于标准规定一级米13%~18%的范围内。另外,福恢676及其他4个恢复系的胶稠度均大于60 mm,达到一级米标准。
表 3 米质分析Table 3. Analysis of grain quality种植地区Growing Areas 恢复系Restorer lines 糙米率Brown rice rate/% 精米率Milled rice rate/% 整精米率Head rice rate/% 长Length/mm 宽Width/mm 长宽比Length to width 垩白率Chalkiness ratio/% 垩白度Chalkiness degree/% 透明度Transparency 碱消值Alkali value 直链淀粉含量Amylose content/% 胶稠度Gel consistency/mm 福州Fuzhou 福恢676 Fuhui676 81.40±0.13 69.60±0.28 44.10±0.59 6.39±0.11 2.23±0.09 2.88±0.02 28.13±0.97 6.23±0.87 3.00±0.00 4.53±0.12 14.20±0.33 91.97±0.29 明恢63 Minghui63 81.80±0.26 69.80±0.96 38.27±1.06* 6.39±0.19 2.30±0.02 2.80±0.04* 18.83±1.30* 3.70±0.50* 3.00±0.00 4.30±0.10 11.97±0.31* 90.97±1.16 蜀恢527 Shuhui527 81.87±0.64 69.30±0.79 35.10±0.56* 6.95±0.25* 2.30±0.06 3.12±0.07* 16.87±0.38* 3.23±0.15* 2.00±0.00 4.43±0.06 12.47±0.12* 86.00±0.80* 明恢86 Minghui86 82.13±0.15* 68.37±0.50* 43.10±0.79 6.19±0.12* 2.32±0.09 2.70±0.06* 35.97±1.22* 9.93±0.38* 2.00±0.00 6.83±0.12* 13.50±0.17* 83.00±0.60* 福恢673 Fuhui673 82.23±0.15* 68.10±0.44* 41.60±0.90* 6.23±0.13 2.29±0.02 2.74±0.01* 49.57±1.76* 10.10±0.26* 3.00±0.00 4.27±0.15 12.17±0.12* 95.00±0.30* 泉州Quanzhou 福恢676 Fuhui676 81.80±0.15 71.23±0.31 55.43±0.90 6.71±0.15 2.27±0.07 2.98±0.07 16.30±0.82 3.60±0.46 2.00±0.00 4.33±0.21 15.40±0.53 88.00±0.61 明恢63 Minghui63 82.80±0.15* 71.37±0.51 40.60±0.30* 6.83±0.10 2.38±0.05 2.90±0.04 14.13±0.76* 2.57±0.31* 3.00±0.00 4.37±0.21 13.00±0.20* 90.03±0.06* 蜀恢527 Shuhui527 81.8±0.17 70.77±0.23 42.67±0.21* 7.23±0.21* 2.27±0.06 3.21±0.04* 11.60±0.61* 1.83±0.31* 2.00±0.00 4.30±0.26 14.23±0.38* 86.00±0.75* 明恢86 Minghui86 82.60±0.15* 70.63±0.15* 36.80±0.17* 6.62±0.15 2.40±0.04* 2.79±0.07* 38.63±0.49* 9.07±0.70* 2.00±0.00 6.47±0.29* 15.16±0.55 78.00±0.72* 福恢673 Fuhui673 81.70±0.20 70.60±0.20* 42.33±0.47* 6.83±0.25 2.34±0.07 2.94±0.05 50.93±3.23* 10.23±0.83* 3.00±0.00 4.20±0.10 13.57±0.21* 93.03±0.21* 三明Sanming 福恢676 Fuhui676 82.70±0.10 73.37±0.21 51.80±0.70 6.63±0.10 2.28±0.05 2.92±0.06 17.9±1.42 3.67±0.31 2.00±0.00 4.43±0.15 16.67±0.32 92.00±0.50 明恢63 Minghui63 81.90±0.06* 71.63±0.15* 44.30±0.44* 6.75±0.19 2.41±0.08 2.81±0.07 9.20±0.56* 1.73±0.49* 2.00±0.00 4.83±0.06* 16.17±0.15 86.03±0.74* 蜀恢527 Shuhui527 81.90±0.35* 72.10±0.40* 34.10±0.44* 7.23±0.21* 2.36±0.05 3.14±0.09* 12.80±2.26* 2.83±0.55 3.00±0.00 4.43±0.32 15.83±0.06* 91.97±1.25 明恢86 Minghui86 83.20±0.26* 74.07±0.23* 38.67±1.26* 6.32±0.12* 2.39±0.10 2.65±0.05* 18.30±1.87 5.43±0.25* 2.00±0.00 6.83±0.21* 16.07±0.51 81.97±0.64* 福恢673 Fuhui673 82.40±0.10* 71.03±0.21* 49.20±0.61* 6.48±0.07* 2.34±0.08 2.79±0.02* 29.73±0.80* 6.53±0.55* 3.00±0.00 4.33±0.25 14.43±0.31* 92.00±0.40 2.4 粒型及米制相关基因表达分析
进一步采用qRT-PCR分析了福恢676、明恢63、蜀恢527、明恢86及福恢673的稻谷灌浆4 d、7 d和11 d时控制粒形及米质相关基因的表达模式(图4、图5)。
2.4.1 粒形控制相关基因的表达分析
GL7是控制粒长和粒宽的主效基因。在福恢676中,灌浆7 d时GL7的表达略低于灌浆4 d时的表达,但变化不明显,而灌浆11 d时表达明显下降。在明恢63和福恢673中,GL7的表达在3个时期内随着灌浆时期的延长逐渐降低。在蜀恢527和明恢86中,GL7在灌浆4 d时表达量最高,灌浆7 d时明显降低,而灌浆11 d时表达又有所提高,尤其是在蜀恢527中。在福恢676和其他4个恢复系中,GL7均在灌浆4 d时表达量最高。
GS3是控制水稻粒重和粒长的主效基因,同时也是控制水稻粒宽和籽粒充实度的微效基因,在调节籽粒和器官大小中发挥负调节子的功能。在福恢676中,GS3的表达在灌浆7 d时比4 d时稍微提高,而灌浆11 d时表达急剧降低;在亲本明恢63和蜀恢527中,灌浆4 d和7 d时GS3的表达没有明显变化,而灌浆11 d时表达明显下降;在明恢86和福恢673中,GS3的表达在灌浆7 d时比4 d时明显降低,但灌浆11 d时又明显回升且明显高于福恢676及亲本明恢63和蜀恢527。另外,灌浆7 d时,福恢676中GS3的表达量均明显高于其他4个恢复系中的表达量。
GW8是同时控制谷粒大小、粒型和稻米品质的基因。在福恢676和明恢63中, GW8的表达先降低后升高,在福恢676中灌浆11 d时表达量最高,而在明恢63中灌浆4 d时表达量较高。在蜀恢527中,灌浆4 d和7 d时GW8的表达没有明显变化,而在灌浆11 d有所提高。在明恢86和福恢673中,GW8的表达量在灌浆4 d时最高,灌浆7 d时急剧下降,灌浆11 d时又稍回升。
SGL调控水稻种子长度。在福恢676和蜀恢527中,SGL的表达先提高后降低,且在灌浆7 d时表达量较高,福恢676中灌浆11 d时SGL的表达急剧降低。在明恢63和明恢86中,SGL的表达逐渐提高,在灌浆11 d时表达量最高。在福恢673中,SGL的表达先降低后升高,且在灌浆11 d时表达量最高。灌浆11 d 时,福恢676中SGL的表达量明显低于其他4个恢复系中的表达量。
GIF1是控制籽粒充实度相关的基因。在福恢676和明恢63中,GIF1的表达先升高后降低,且在灌浆7 d时表达量最高。在蜀恢527、明恢86和福恢673中,灌浆4 d、7 d和11 d的 GIF1表达量逐渐降低,尤其是在福恢673中降低明显。
2.4.2 米质控制相关基因的表达分析
Chalk5是影响水稻籽粒垩白的形成和精米率等的基因。在福恢676和明恢63中,Chalk5的表达量在灌浆7 d时较4 d时升高,而在灌浆11 d时几乎不表达。在蜀恢527中,Chalk5的表达量逐渐降低,在灌浆11 d时表达量极低。明恢86和福恢673中,Chalk5的表达量逐渐降低,在福恢673中降低明显,但在灌浆11 d时仍有少量表达。
ALK控制稻米的糊化温度,并与垩白、直链淀粉含量等相关。ALK的表达模式在福恢676中与在其他4个恢复系中不同,在福恢676中ALK的表达先升高后降低,且在灌浆7 d时表达量最高。在明恢63、蜀恢527和明恢86中,ALK的表达逐渐上升,且在灌浆11 d时表达量较高。在福恢673中,灌浆7 d时ALK的表达稍低于4 d时的表达但变化不明显,而在灌浆11 d时表达量明显提高。灌浆11 d时,福恢676中ALK 的表达量明显低于其他4个恢复系中的表达量。
OsSSI影响水稻籽粒中支链淀粉的结构,从而影响稻米的糊化温度。OsSSI的表达模式在福恢676中与在其他4个恢复系中完全不同,在福恢676中OsSSI的表达先升高后降低,且在灌浆7 d时表达量最高;而在明恢63、蜀恢527、明恢86和福恢673中,OsSSI的表达逐渐上升,且在灌浆11 d时表达量最高。灌浆11 d时,福恢676中OsSSI的表达量明显低于其他4个恢复系中的表达量。
OsBEIIb影响水稻胚乳中淀粉的结构,并与垩白等性状相关。OsBEIIb的表达模式在福恢676中与在其他4个恢复系中也完全不同,在福恢676中OsBEIIb的表达先升高后降低,且在灌浆7 d时表达量最高;而在其他4个恢复系中,OsBEIIb的表达逐渐提高,且在灌浆11 d时表达量最高。灌浆11 d时,福恢676中OsBEIIb的表达量明显低于其他4个恢复系中的表达量。
3. 讨论
粒形是影响水稻产量和稻米品质的重要因素之一,已成为水稻高产优质分子设计育种的重要靶标性状[7, 26]。福恢676的亲本之一蜀恢527是重穗型籼稻恢复系[27],长粒,长宽比大。本研究分析显示,福恢676的谷粒长和长宽比也均大于亲本明恢63及生产上常用的其他两个恢复系明恢86和福恢673;同时福恢676的千粒重也大于明恢63、明恢86和福恢673,极大可能是受到了蜀恢527的遗传因素影响。
如今,水稻育种在注重提高产量的同时也越来越重视稻米品质的提升。稻米品质包括加工品质、外观品质和蒸煮品质,加工品质主要包括糙米率、精米率、整精米率,外观品质主要包括粒形、透明度、垩白粒率、垩白度等,蒸煮品质主要括糊化温度、直链淀粉含量、胶稠度等。本研究结果表明福恢676及其余4个恢复系都具有较高的糙米率,均达81% 以上。值得特别注意的是,种植于3个不同地区的福恢676的整精米率均高于蜀恢527、明恢63、明恢86和福恢673;周勇等[15]研究表明提高恢复系整精米率是突破杂交水稻整精米率偏低的关键。福恢676具较高的整精米率,值得进一步研究分析和应用。福恢676的垩白度和垩白率均高于亲本明恢63和蜀恢527,某种程度上说明相关遗传调控方面较为复杂;同时,种植于泉州和三明的福恢676的垩白度和垩白率较种植于福州的低,相应的透明度也较低,说明这些指标在一定程度上受到了环境的影响。直链淀粉含量是优质米的重要标准,相关研究表明在非糯品种中Wx的基因型籼稻以Wxa 为主,粳稻基本为Wxb;本研究中籼型恢复系福恢676的Wx基因型与亲本蜀恢527和明恢63的一致,均为Wxb,是因为它们具有粳稻亲缘成分。种植于3个不同地区的福恢676的直链淀粉含量均处于标准规定一级米范围内(13%~18%),即与具有较低直链淀粉含量的基因型Wxb一致。
稻谷粒形及稻米品质均受相关基因的调控。相关研究表明GL7的表达量提高会促进细胞的延长[10]。本研究qRT-PCR分析表明GL7在福恢676和其他4个恢复系中均在灌浆4 d时表达量最高,说明GL7很可能在灌浆前期发挥较重要的作用。 GS3对籽粒大小和重量起负调控作用[11],在灌浆11 d 时GS3在福恢676及其亲本明恢63、蜀恢527中表达水平较低,但在明恢86和福恢673中表达水平较高,尤其是在明恢86中表达水平最高;稻谷粒形分析显示与福恢676及其亲本明恢63、蜀恢527相比,3个种植地区的明恢86和福恢673的谷粒均较短,其中明恢86的谷粒最短,在千粒重方面,除种植于泉州的福恢673外,明恢86和福恢673的千粒重也均较低;因此,本研究中稻谷粒长与千粒重的表型分析结果与GS3的表达水平一致。GIF1控制灌浆并与籽粒的充实度有关[14]。本研究分析显示灌浆11 d时GIF1的表达量在5个恢复系中均明显降低,应该是由于11 d时已经是灌浆后期,因此基因表达也相应发生了变化。另外,福恢676的控制米质相关基因ALK、OsSSI、OsBEIIb的表达模式均与其他4个恢复系的完全不同,很可能是引起整精米率等有差异的原因。但关于相关基因如何调控影响福恢676的粒形和米质尤其是粒长和整精米率等方面有待进一步的分析和研究。
4. 结论
本研究分析表明恢复系福恢676的粒长较长,长宽比相对较大,整精米率较高,糙米率、胶稠度和直链淀粉含量达到一级米标准;Wx的基因型为Wxb,福恢676中控制粒形基因GL7、GS3和控制米质基因ALK 、 OsSSI、OsBEIIb的表达模式与其他恢复系中有明显不同,为福恢676应用于育种生产及解析其杂交后代的遗传基础方面提供理论依据。
-
![]()
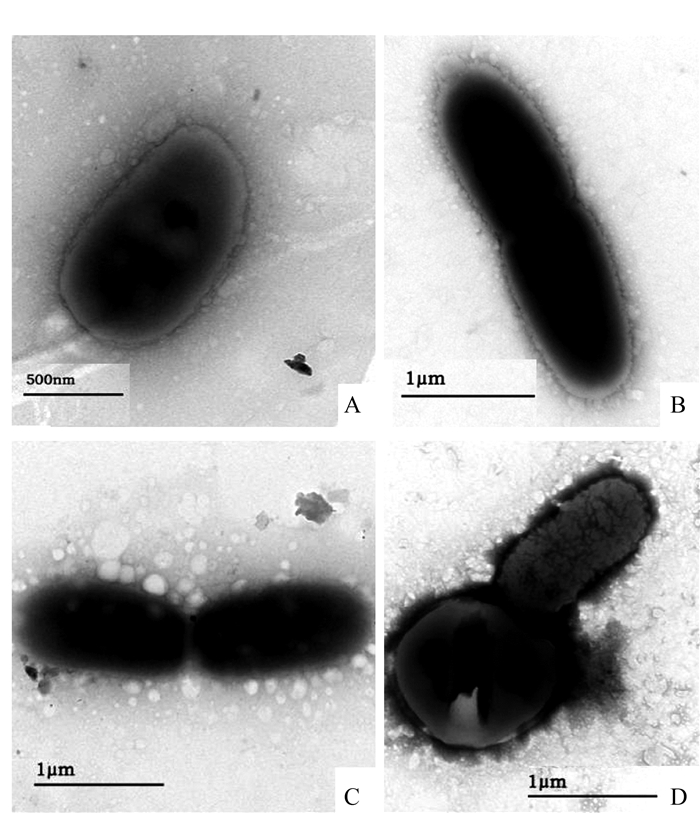
图 1 菌株FJAT-1458发酵过程菌体形态变化
Figure 1. Morphological changes on FJAT-1458 during fermentation process
表 1 植物疫苗菌株FJAT-1458在不同培养条件下的生长量
Table 1 Growth of avirulent strain, FJAT-1458, under varied culture conditions
测定项目 FJAT-1458菌体浓度/
(×108cfu·mL-1)培养基 SPA 1.56±0.12 a NA 0.93±0.05 c LB 1.39±0.11 a 温度 20℃ 1.14±0.06 bc 25℃ 1.27±0.12 b 30℃ 1.77±0.10 a 35℃ 0.91±0.06 c 40℃ 0 装液量 15mL 1.82±0.07 a 25 mL 1.75±0.07 a 35 mL 1.87±0.11a 45 mL 1.77±0.04 a 55 mL 1.71±0.06a 注:同例不同小写字母表示差异达显著水平(P < 0.05)。表 2同。  下载: 导出CSV
下载: 导出CSV
表 2 植物疫苗菌株FJAT-1458发酵过程各参数变化
Table 2 Changes on parameters during fermentationprocess of avirulent strain, FJAT-1458
时间
/hFJAT-1458菌体浓度
/(×108cfu·mL-1)菌株弱
化指数pH值 0 1.12±0.07 a 0.84±0.03 a 7.23±0.01 h 6 1.37±0.15 a 0.84±0.01 a 7.51±0.01 g 12 3.05±1.56 b 0.84±0.03 a 8.27±0.01 f 24 9.37±1.02 c 0.85±0.01 a 8.53±0.03 d 36 12.48±2.55 d 0.85±0.01 a 8.38±0.02 e 48 11.72±1.82 e 0.85±0.01 a 8.72±0.01 b 60 11.05±1.22 f 0.85±0.02 a 8.67±0.01 c 72 8.32±0.43 g 0.83±0.03 a 8.78±0.03 a
下载: 导出CSV
-
[1] HAYWARD A C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum [J]. Annual Review of Phytopathology, 1991, 29: 65-87. DOI: 10.1146/annurev.py.29.090191.000433
[2] VANITHA S C, NIRANJANASR, UMESHA S. Role of phenylalanine ammonia lyase and polyphenol oxidase in host resistance to bacterial wilt of tomato [J]. Journal of Phytopathology, 2009, 157(9): 552-557. DOI: 10.1111/jph.2009.157.issue-9
[3] 黄真池, 刘媛, 曾富华, 等.青枯菌致病机理及作物抗青枯病研究进展[J].热带亚热带植物学报, 2008, 16(5): 491-496. http://www.cnki.com.cn/Article/CJFDTOTAL-RYZB200805016.htm [4] 袁立和, 苏清岚, 易由华.番茄青枯病防治技术研究[J].中国蔬菜, 1992, 18(5): 4-6. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGSC199205001.htm [5] JONES J D G, DANGL J L. The plant immune system[J]. Nature, 2006, 444(16): 323-329. http://cn.bing.com/academic/profile?id=2162515800&encoded=0&v=paper_preview&mkt=zh-cn
[6] CHANG Y H, YAN H Z, LIOU R F. A novel elicitor protein from phytophthora parasitica induces plant basa immunity and systemic acquired resistance [J]. Molecular Plant Pathology, 2015, 16(2):123-136. DOI: 10.1111/mpp.2015.16.issue-2
[7] DALE W, ADRIAN N, GARY L. Induced resistance for plant defence [M]. Oxford: Blackwell Publishing, 2007: 31-81.
[8] 邱德文.植物免疫与植物疫苗研究与实践[M].北京:科学出版社, 2008. [9] NORMAN D J, ZAPATA M, GABRIEL D W, et al. Genetic diversity and host range variation of Ralstonia solancearum strains entering North America [J]. Phytopathology, 2009, 99(9): 1070-1077. DOI: 10.1094/PHYTO-99-9-1070
[10] CELLIER G, PRIOR P. Deciphering phenotypic diversity of Ralstonia solanacearum strains pathogenic to potato [J]. Phytopatholy, 2010, 100: 1250-1261. DOI: 10.1094/PHYTO-02-10-0059
[11] SUGA Y, HORITA M, UMEKITA M, et al. Pathogenic characters of Japanese potato strains of Ralstonia solanacearum [J]. Journal of General Plant Pathology, 2013, 79: 110-114. DOI: 10.1007/s10327-013-0429-7
[12] 刘波, 蓝江林, 朱育菁, 等.植物免疫系统的研究与应用[J].中国农学通报, 2007, (23): 163-172. http://wiki.cnki.com.cn/HotWord/6090486.htm [13] 郑雪芳, 刘波, 林乃铨, 等.青枯雷尔氏菌无致病力突变菌株的构建及其防效评价模型分析[J].植物病理学报, 2013, 43(5): 518-531. http://www.cnki.com.cn/Article/CJFDTOTAL-ZWBL201305010.htm [14] KELMAN A. The relationship of pathogenicity in P. solanacearum to colony appearance on a tetrazolium medium [J]. Phytopathology, 1954, 44: 693-695. https://www.cabdirect.org/cabdirect/abstract/19551101405
[15] 刘波, 林营志, 朱育菁, 等.生防菌对青枯雷尔氏菌的致弱特性[J].农业生物技术学报. 2004, 12(3): 322-329. http://www.cnki.com.cn/Article/CJFDTOTAL-NYSB200403015.htm [16] 陈燕萍, 刘波, 葛慈斌, 等.作物青枯病植物疫苗工程菌FJAT-T8发酵过程生长适合度研究[J].福建农业学报, 2012, 27(3): 287-293. http://www.fjnyxb.cn/CN/abstract/abstract1874.shtml [17] 周青, 王笑, 苏婷, 等.青枯病和根肿病生防细菌BS2004的发酵培养基和培养条件优化[J].中国生物防治学报, 2012, 28(4): 537-544. http://www.cnki.com.cn/Article/CJFDTOTAL-ZSWF201204014.htm [18] 马耀华, 谭小艳, 黄思良, 等.石榴干腐病生防菌株Z2的鉴定及其培养条件的优化[J].植物病理学报, 2015, 45(4): 425-437. http://www.cnki.com.cn/Article/CJFDTOTAL-ZWBL201504012.htm [19] 巫建新, 席刚俊, 杨卫君, 等.抑制杨木变色生防细菌的筛选及其发酵培养条件优化研究[J].西部林业科学, 2013, 42(2): 99-102. http://www.cnki.com.cn/Article/CJFDTOTAL-YNLK201302018.htm [20] 车建美, 蓝江林, 刘波.转绿色荧光蛋白基因的青枯雷尔氏菌生物学特性[J].中国农业科学, 2008, 41(11): 3626-3635. http://www.cnki.com.cn/Article/CJFDTOTAL-ZNYK200811026.htm [21] EINSTONE J G, HENNESSY J, WILSON J K, et al Sensitivity of different methods for the detection of Ralstonia solanacearum in potato tuber extracts. EPPO Bull, 1996, 26: 674-678. http://cn.bing.com/academic/profile?id=2006967948&encoded=0&v=paper_preview&mkt=zh-cn
-
期刊类型引用(1)
1. 赵培栋,舒添,刘沛,张振乾,杨柳. 油菜未成熟种子含水量近红外快速测定方法. 作物研究. 2025(01): 81-87 .  百度学术
百度学术
其他类型引用(1)





计量
- 文章访问数: 1398
- HTML全文浏览量: 153
- PDF下载量: 264
- 被引次数: 2
