

Inheritance of a Growth Characteristic of Precocious Restorer Rice Line, DR02
-
摘要: 水稻DR02来源于T251与02K511的杂交后代,具有显性早熟特性。以DR02分别与明恢86、明恢63等6个不同类型的中、迟熟水稻品种杂交后代进行遗传性分析。结果显示,各组合F1的抽穗期均比中亲值早,DR02与双抗明占和9311的杂交F2的抽穗期呈双峰分布,与其他4个亲本的F2抽穗期呈单峰分布,用中亲值进行分组,经χ2测验,仅DR02与粤晶丝苗2号和双抗明占组合的F2代分离比符合3:1,其他组合均表现显著与极显著差异,不符合预期理论比。综合上述分析结果表明水稻材料DR02的早熟性可能是由1个比较大效应的主效QTL控制的。Abstract: A dominant early maturing trait was identified in DR02 rice bred from T251 and 02K511. Heading dates of the hybrid progenies from DR02 crossed with 6 medium or late maturing varieties, including Minghui 86 and Minghui 63, were analyzed to study the inheritance of the growth characteristic. It was found that the heading dates of F1 progenies were earlier than those of their mid-parents. The heading dates of F2 populations from DR02 crossed with Shuangkangmingzhan and 9311 had twin peaks, while those of F2 from DR02 crossed with the other 4 parents exhibited a unimodal distribution with closely clustered values. The segregation ratio between the early heading and the late heading plants was 3:1 for the F2 population of DR02 crossed with Yuejingsimiao 2 and Shuangkangmingzhan, while those of other combinations were significantly and extremely significantly different. It suggested that the early maturing trait of DR02 could be largely controlled by a single relatively dominant QTL.
-
Keywords:
- rice /
- early maturing /
- restorer line DR02
-
羊的传统放牧方式受到天然草场营养物质的季节性不平衡、载畜量有限以及草地生态环境恶化等因素的制约[1],舍饲圈养是我国未来肉羊饲养方式的发展方向[2-3]。有研究者比较了不同饲养方式对南江黄羊[3]、浏阳黑山羊[4]等肉羊品种生长发育的影响。福清山羊是福建省优良地方肉用品种,具有早熟、繁殖力高、耐粗饲、肉质风味好等优点,深受消费者喜爱[5]。谢喜平等研究了福清山羊和成都麻羊F1代早期生长发育规律[6],李文杨等采用生长曲线拟合法研究了放牧福清山羊体重生长发育规律[7],但尚未见不同饲养方式对福清山羊生长发育影响的报道。为了深入保护和开发福清山羊种质资源,拟比较放牧和舍饲圈养模式下福清山羊羯羊的生长发育规律,并利用Von Bertalalanffy、Gompertz和Logistic非线性生长模型对不同饲养方式的福清山羊羯羊体重增长进行拟合分析,为福清山羊商品羊的饲养管理和生产模式优化提供理论依据。
1. 材料与方法
1.1 试验材料与饲养管理
试验在福建省农业科学院福清市渔溪肉羊试验基地进行。采用单因素试验设计,选择体重、体况相近的健康3月龄断奶福清山羊(2017年1-2月出生)羯羊90只,平均体重(9.33±1.34) kg。按照饲养方式分为全圈养组(A组)、圈养+放牧组(B组)、放牧组(C组)3组,每组30只,各组在相同面积的羊栏(14.0 m×6.3 m)内单栏饲养。其中A组不放牧(全圈养饲养),B组和C组在非雨天进行每天4~7 h的相同时长放牧,B组的放牧地为固定的15 m×30 m舍外草地(圈养结合放牧饲养),C组天然草场人工放牧(放牧饲养)。试验开始前对试验羊只进行体重、体尺测定和统一编号。
试验日粮参考肉羊饲养标准(NY/T816-2004)(日增重为100 g·d-1)按照4~6月龄(前期)、7~9月龄(中期)、10~12月龄(后期)3个时期分别设计。A组、B组饲喂日粮、日饲喂量相同,前、中、后期饲喂量(干物质计)分别为0.56、0.61、0.66 kg·d-1;C组放牧后进行补饲,补饲日粮和补饲量与A组日粮中精料配方一致;早晚各饲喂(补饲)1次。不同时期各日粮组成及营养水平见表 1。试验期间各组羊只自由采食和饮水,栏舍环境条件及其他饲养管理均相同。
表 1 试验日粮组成及营养水平(干物质基础)Table 1. Formulation and nutritional composition of experimental diet (DM basis)项目 4~6月龄 7~9月龄 10~12月龄 A、B组 C组 A、B组 C组 A、B组 C组 精料 牧草 精料 牧草 精料 牧草 原料 天然牧草 100 100 100 玉米青贮/% 30 30 30 杂交狼尾草/% 30 30 30 玉米/% 18 45 25 62.5 23 57.5 大豆粕/% 10.3 25.75 4 10 2.5 6.25 麸皮/% 8.2 20.5 7.4 18.5 11 27.5 磷酸氢钙/% 1.1 2.75 1.2 3 1.15 2.875 食盐/% 0.5 1.25 0.5 1.25 0.5 1.25 小苏打/% 0.5 1.25 0.55 1.375 0.5 1.25 石粉/% 0.9 2.25 0.85 2.125 0.85 2.125 预混料①/% 0.5 1.25 0.5 1.25 0.5 1.25 合计/% 100 100 100 100 100 100 100 100 100 营养水平 消化能②/(MJ·kg-1) 6.39 12.52 6.39 12.50 6.33 12.35 粗蛋白/% 13.21 18.02 7.57 10.67 12.45 10.24 10.44 11.75 6.14 钙/% 0.86 1.79 0.98 0.79 1.72 0.95 0.74 1.66 1.02 磷/% 0.57 1.02 0.09 0.49 1.08 0.18 0.50 1.11 0.11 中性洗涤纤维/% 47.28 29.11 66.77 52.77 37.69 61.33 56.71 38.11 67.48 酸性洗涤纤维/% 27.67 11.75 41.97 29.61 13.94 39.66 31.31 13.46 42.15 注:①每千克预混料中含有:VA 200000 IU,VD 50000 IU,VE 500 IU,Fe 2 g,Cu 0.75 g,Zn 3 g,Mn 4 g,I 50 mg,Se 20 mg,Co 50 mg。②除消化能为计算值外,其他均为实测值。 1.2 体重测定
试验全期9个月,于每个时期的前1 d以及试验结束当天上午逐只空腹称重。
1.3 拟合分析模型
选用Von Bertalanffy、Gompertz和Logistic 3种数学模型作为非线性生长模型,对不同饲养方式福清山羊体重生长过程进行拟合。各模型中W表示t月龄时的体重估计值,参数A为成熟体重,K为瞬时相对生长率,B为调节参数,t为月龄(表 2)。
表 2 拟合分析的3种非线性生长模型表达式及特征参数Table 2. Three nonlinear models and parameters for analyzing growth of Fuqing goats模型 表达式 拐点体重 拐点月龄 Von Bertalanffy W = A(1- Be-Kt)3 8A/27 (ln3B)/K Gompertz W = Ae-bexp(-Kt) A/e (lnB)/K Logistic W = A/(1+Be-Kt) A/2 (lnB)/K 1.4 数据处理及分析方法
参考戴国俊等[8]的方法,利用SPSS17.0软件进行Von Bertalanffy、Gompertz和Logistic 3种非线性生长模型分析,并估计各模型的A、B、
K参数和拟合度R2,进一步建立各生长曲线模型。基础数据统计计算采用Excel 2007。 2. 结果与分析
2.1 不同饲养方式下福清山羊生长情况
不同饲养方式下,相同月龄放牧组试验羊的体重、平均日增重和相对生长率均为最低(表 3)。3种饲养方式试验羊的6月龄体重差异不显著(P>0.05),A组体重最大,为15.83 kg。B组试验羊9月龄和12月龄的体重显著高于放牧组(P<0.05),分别为20.95和26.03 kg;A组2个阶段的体重高于C组,二者差异不显著水平(P>0.05)。
表 3 不同饲养方式福清山羊体重增长的影响Table 3. Effect of husbandry methods on body weight of Fuqing goats月龄 饲养方式 体重/kg 平均日增重/(g·d-1) 相对生长率/% 3 A组 9.32±1.37 - - B组 9.33±1.35 - - C组 9.34±1.35 - - 6 A组 15.98±1.79 73.96±6.40a 53.00±4.20a B组 15.83±1.81 72.30±5.90a 52.05±3.35a C组 15.39±1.81 67.30±5.82b 49.30±3.40b 9 A组 20.67±2.88ab 51.19±12.61b 25.28±3.26b B组 20.95±2.09a 56.89±5.74a 27.99±3.01a C组 19.60±2.23b 46.70±6.23c 24.07±2.13b 12 A组 25.09±4.60ab 49.04±20.51ab 18.48±6.10ab B组 26.03±3.10a 56.44±13.74a 21.35±4.36a C组 23.62±3.81b 44.67±19.38b 17.89±6.54b 注:同列数据后不同小写字母表示相同月龄之间差异显著(P<0.05)。 随着日龄的增长,各饲养方式的平均日增重和相对生长率均呈下降趋势(表 3),符合畜禽体重生长发育的一般规律。整个试验周期内,3~6月龄福清山羊羯羊的生长速度最快,其中A组平均日增重和相对生长率最大,分别为73.96 g·d-1、53.00%,A组和B组平均日增重和相对生长率显著高于C组(P<0.05)。6~12月龄,B组平均日增重和相对生长率最大,均显著高于C组(P<0.05),6~9月龄时显著高于A组(P<0.05),9~12月龄与A组差异不显著;C组全期平均日增重和相对生长率均最小。
2.2 不同饲养方式下福清山羊生长模型拟合分析
不同饲养方式下福清山羊生长模型拟合结果见表 4。拟合度(R2)是评价非线性生长模型拟合实际生长发育的主要指标,R2越接近1说明拟合效果越好。由表 4可知,3种模型的拟合度均较高(R2>0.99),表明选用的生长模型都能很好地描述不同饲养方式福清山羊羯羊的体重生长发育趋势与规律,其中Von Bertalanffy模型的拟合度最高,该模型A组、B组和C组福清山羊羯羊的拟合度分别高达0.999、0.999和0.998(表 4)。通过进一步利用3种数学模型计算各试验组不同时期的体重理论值(表 5),Von Bertalanffy模型计算的理论值更接近实际值,也证实该模型的拟合效果最优。
表 4 不同饲养方式下3种模型拟合参数Table 4. Fitting parameters of three models for growth patterns of Fuqing goats raised under different husbandry methods饲养方式 生长模型 A B K R2 拐点月龄 拐点体重/kg A组 Von Bertalanffy 33.583 0.533 0.145 0.999 3.24 9.95 Gompertz 31.711 2.079 0.180 0.998 4.06 11.67 Logistic 28.684 4.757 0.287 0.996 5.43 14.34 B组 Von Bertalanffy 39.960 0.542 0.116 0.999 4.19 11.84 Gompertz 36.476 2.136 0.153 0.999 4.96 13.42 Logistic 31.491 5.059 0.263 0.997 6.16 15.75 C组 Von Bertalanffy 31.564 0.505 0.141 0.998 2.95 9.35 Gompertz 29.894 1.940 0.174 0.998 3.81 11.00 Logistic 27.130 4.212 0.274 0.996 5.25 13.57 表 5 不同饲养方式下3种生长曲线模型体重估计值与实际值Table 5. Observed and model-estimated body weights of Fuqing goats raised under different husbandry methods饲养方式 生长模型 体重/kg 平均日增重/(g·d-1) 相对生长率/% 3月龄 6月龄 9月龄 12月龄 3~6月龄 6~9月龄 9~12月龄 3~6月龄 6~9月龄 9~12月龄 A组 实际值 9.32 15.98 20.67 25.09 74.0 52.1 49.1 52.6 25.6 19.3 Von Bertalanffy 9.44 15.74 21.02 25.01 70.0 58.7 44.3 50.0 28.7 17.3 Gompertz 9.44 15.65 21.01 24.95 69.0 59.6 43.8 49.5 29.2 17.1 Logistic 9.53 15.50 21.10 24.90 72.2 56.9 56.4 51.7 27.8 21.6 B组 实际值 9.33 15.83 20.95 26.03 68.1 62.7 52.4 49.2 30.7 20.1 Von Bertalanffy 9.40 15.53 21.17 25.89 67.6 63.8 51.9 48.6 31.2 19.8 Gompertz 9.46 15.54 21.28 25.95 67.2 46.8 44.7 48.9 24.1 18.6 Logistic 9.55 15.41 21.36 25.91 63.4 53 40.1 46.4 27.2 16.6 C组 实际值 9.34 15.39 19.60 23.62 62.7 53.7 39.8 45.9 27.6 16.5 Von Bertalanffy 9.46 15.17 19.94 23.55 74.0 52.1 49.1 52.6 25.6 19.3 Gompertz 9.46 15.10 19.93 23.51 70.0 58.7 44.3 50.0 28.7 17.3 Logistic 9.51 14.96 19.98 23.44 69.0 59.6 43.8 49.5 29.2 17.1 利用Von Bertalanffy模型预测A组、B组和C组的成熟体重分别为33.583、39.96和31.564 kg,拐点月龄分别为3.24、4.19和2.95,拐点体重分别为9.95、11.84和9.35 kg。同一生长模型拟合不同饲养方式的成熟体重、拐点月龄和拐点体重均呈现相同规律,为C组<A组<B组(表 3)。表明采用B组的饲养方式对福清山羊羯羊的生长发育具有促进作用,主要通过生长拐点月龄的延迟,以及拐点体重和成熟体重的增加实现。
3. 讨论
3.1 不同饲养方式对福清山羊生长发育的影响
羊只的不同饲养方式育肥效果存在差异的主要原因是摄取的营养物质不同[9]。苏铁等[4]、温琦等[10]研究表明,对放牧羔羊进行补饲可提高育肥速度。李文杨等[7]研究了放牧福清山羊生长发育情况,表明周岁内(2~12月龄)福清山羊的平均日增重为51.4 g·d-1,低于本研究的3个试验组(52.9~61.9 g·d-1),精料补饲水平的差异是造成福清山羊日增重差异的关键原因。另外,虽然本研究的试验日粮参考NRC标准,设计日增重为100 g·d-1的营养摄入水平,但3个试验组的日增重远低于预期,表明遗传背景也是影响肉羊生长发育的重要制约因素。
在日粮中补饲相同营养水平的精料,圈养福清山羊羯羊的增重效果优于放牧组,与谢喜平等[3]、武原红等[9]研究结果一致,这一方面是由于放牧条件的粗饲料来源不确定和不稳定造成的[3],另一方面不同饲养方式羊只的运动和防寒耗能不同也影响了增重效果[9]。张明等[11]研究表明,在相同日粮水平圈养下,降低饲养密度能提高小尾寒羊的生产性能;王磊等[12]也证实了圈养饲养面积越大肉羊增质量效果越好。饲喂相同日粮的前提下,B组6~12月龄的日增重高于A组,初步表明圈养福清山羊活动面积的增加会促进其生长。
3.2 不同饲养方式福清山羊生长模型拟合分析
Von Bertalanffy、Gompertz 和Logistic 3种非线性生长模型是拟合畜禽生长发育的常用的数学模型,3种模型对各试验组的拟合度均在0.99以上,拟合效果较理想,Von Bertalanffy模型的拟合效果最优。羊的生长发育遵循一定的规律,但又受到品种、生产目的、饲养管理方式等多种因素影响,不同品种间最优非线性模型也存在差异[13]。大量的研究表明Gompertz和Von Bertalanffy 模型更适合羊的生长发育拟合分析[7, 13-15],可较准确地描述大部分羊品种的生长发育规律。李文杨等[7]研究了放牧福清山羊的生长发育规律,表明Von Bertalanffy 模型对福清山羊公、母羊的拟合度高于其他模型,拟合效果最优,与本研究结果一致。
畜禽体重生长曲线是改进其饲养管理方式以及实现商品畜禽适时出栏的主要依据。采用拟合效果最优的Von Bertalanffy模型预测各试验组的成熟体重、瞬时相对生长率、拐点体重和拐点月龄均存在差异,表明饲养方式是福清山羊商品羊育肥的主要制约因素之一。B组育肥效果最优的主要原因是其生长拐点的延迟,前期快速增长时间长,而生长拐点后适当降低精料水平可减少饲料成本的支出。在饲喂日粮营养水平相似的条件下,福清山羊商品羊6月龄前采用限制活动面积进行圈养育肥,6月龄后扩大活动面积圈养育肥,体重达25 kg左右上市,可获得较理想的育肥效果和经济效益。
-
![]()
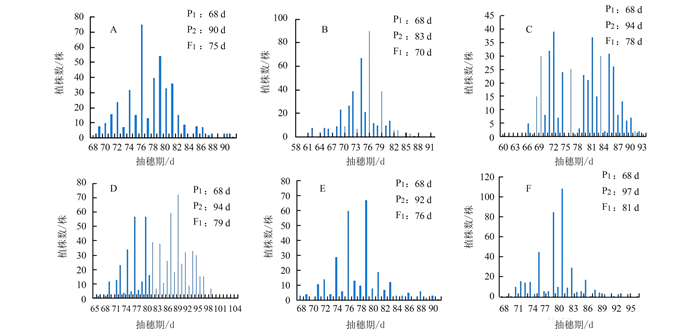
图 1 DR02分别与不同类型品种杂交F2的抽穗期分布
注:A为DR02/成恢727的F2;B为DR02/粤晶丝苗2号的F2;C为DR02/9311的F2;D为DR02/双抗明占的F2;E为DR02/明恢86的F2;F为DR02/明恢63的F2。P1为DR02,P2为中、迟熟亲本,F1为P1与P2的杂交后代。
Figure 1. Distribution of heading dates in F2 populations of DR02 crossed with different types of rice
表 1 DR02与不同类型中、迟熟品种杂交F1的抽穗期
Table 1 Heading dates of F1 progenies from DR02 crossed with various types of medium or late maturing varieties

 下载: 导出CSV
下载: 导出CSV
-
[1] 熊振民, 蔡洪法.中国水稻[M].北京:中国农业科学技术出版社, 1992:43-44. [2] KINOSHITA T. Report of the committee on gene symbolization, nomenclature and linkage groups[J].Rice Genet Newsl, 1995, 12:9-153. DOI: 10.1142/9789812814289_bmatter
[3] ICHITANI K, OKUMOTO Y, TANISAKA T. Genetic analyses of low photo period sensitivity of rice cultivars from the northern most regions of Japan[J]. Plant Breed, 1998, 117:543-547. DOI: 10.1111/pbr.1998.117.issue-6
[4] 沈波, 钱惠荣, 王建林, 等.应用RFLP定位水稻的生育期基因[J].作物学报, 1994, 10(1):1-7. http://www.cnki.com.cn/Article/CJFDTOTAL-XBZW199401000.htm [5] OKUMOTO Y, TANISAKA T, YAMAGATA H. Heading-time genes of the rice varieties grown in the Tohoku-Hokuriku region in Japan[J]. Japanese Journal of Breeding, 1992, 42:121-135. DOI: 10.1270/jsbbs1951.42.121
[6] DUNG L V, SANO Y. Dissection of a gene complex responsible for photoperiod sensitivity[J].Rice Genet News, 1996, 13:72-74.
[7] TSAI K H. Gene loci and alleles controlling the duration of basic vegetative growth of rice[J].Rice Genet Philippines:IRRI, 1986, (1):339-349. DOI: 10.1142/9789812814265_0030?queryID=%24%7BresultBean.queryID%7D
[8] 邓晓建, 周开达, 李仁端, 等.水稻完全显性早熟性的发现和基因定位[J].中国农业科学, 2001, 34(3):233-239. http://www.cnki.com.cn/Article/CJFDTOTAL-ZNYK200103000.htm [9] 李仕贵, 马玉清, 王文明, 等.一个新的水稻迟熟性基因的遗传分析和分子标记定位[J].遗传学报, 2000, 27(2):133-138. http://www.cnki.com.cn/Article/CJFDTOTAL-YCXB200002006.htm [10] 蒋佩琪, 淳泽, 邓晓建, 等.水稻早熟基因显性抑制基因的遗传分析和分子标记定位[J].四川大学学报:自然科学版, 2003, 40(2):377-381. http://cdmd.cnki.com.cn/Article/CDMD-10610-2003106892.htm [11] YANG Y J, WANG X D, W X J, et al. The discovery, genetic analysis and gene mapping of earliness rice(Oryza Sativa L.)D64B[J]. Acta Genet Sin, 2005, 32(5):489-500. https://www.researchgate.net/publication/7726882_The_discovery_genetic_analysis_and_gene_mapping_of_earliness_rice_Oryza_saliva_L_D64B
[12] TSAI K H. A gene, w-Ef-1(t), for weakening the effect of early-heading[J]. Revue Generale Nucleaire, 1996, 13:78-80.
[13] SATO S, OGATA K, SHINJYO C. Thermo-sensitive action of an earliness gene Ef-x in rice, Oryza sativa L.[J]. Japanese Journal of Genetics, 1992, 67:473-482. DOI: 10.1266/jjg.67.473
[14] SATO S, SAKAMOTO I, SHIRAKAWA K. Chromosomal location of an earliness gene Ef-1 of rice, Oryza sativa L.[J].Jpn J Breed, 1988, 38:385-396. DOI: 10.1270/jsbbs1951.38.385
[15] OKUNOTO Y, TANISAKA T. Trisomic analysis of a strong photoperiod-sensitivity gene E1 in rice(Oryza sativa L.)[J].Euphytica, 1997, 95:301-307. DOI: 10.1023/A:1003052400082
-
期刊类型引用(4)
1. 徐倩,吴贤锋,刘远,李文杨. 饲粮中添加不同水平大蒜皮对福清山羊生长性能、营养物质表观消化率、瘤胃发酵参数和微生物结构的影响. 动物营养学报. 2024(10): 6479-6493 .  百度学术
百度学术
2. 李桂贤. 南江黄羊生长曲线拟合分析. 养殖与饲料. 2023(01): 23-27 . 百度学术
3. 林云琴. 福清山羊生产现状及发展对策. 中国畜禽种业. 2022(12): 112-114 . 百度学术
4. 刘远,李文杨,吴贤锋,毛坤明,林云琴,黄勤楼. 福清山羊快长品系的世代选育及生产性能测定分析. 福建农业学报. 2021(03): 305-311 . 本站查看
其他类型引用(2)

计量
- 文章访问数: 1296
- HTML全文浏览量: 142
- PDF下载量: 158
- 被引次数: 6
