

Effect of Humidity on Life of Experimental Lasioderma serricorne (Fabricius)
-
摘要: 在(32±1)℃,光周期为14 L:10 D条件下,通过组建烟草甲实验种群两性生命表的方法,探讨了不同湿度对烟草甲Lasioderma serricorne(Fabricius)生长发育和繁殖的影响。结果表明:在50% RH和90% RH条件下,烟草甲未能完成一个世代的发育,90% RH条件下幼虫在1龄全部死亡,50% RH条件下幼虫在2龄全部死亡;而在60% RH、70% RH和80% RH条件下烟草甲均可完成世代发育。卵期长短除70% RH和90% RH外,其余湿度条件下均无明显差异。幼虫发育期在60% RH和70% RH条件下无明显差异;而在80% RH下1、2、3龄发育时期均为最低,分别为8.08 d、6.82 d和7.05 d;60% RH条件下,烟草甲雌成虫寿命最长为25.84 d,80% RH下雄成虫寿命最长为15.25 d;雌成虫产卵前期(2.55 d)、总产卵前期(58.41 d)和产卵期(7.00 d)在80% RH条件下均为最短,60% RH和70% RH条件下无显著差异。60% RH、70% RH和80% RH等3个湿度条件下,繁殖力、特定年龄-龄期阶段的生命期望值(exj)、特定年龄-龄期阶段的生殖价(Vxj)有显著性差异,其中在60% RH条件下均为最大;随着湿度的增加,净增殖率R0逐渐减小分别为37.69、30.65、15.02,内禀增长率r和周限增长率λ分别为0.05、0.05、0.04和1.05、1.05、1.04,均无显著差异;60% RH和70% RH条件下的世代周期T(分别为72.68 d和72.79 d)无显著差异,但均显著长于80% RH(62.80 d);对烟草甲种群进行预测,结果表明60% RH、70% RH和80% RH等3个湿度条件下烟草甲种群最低虫口数分别出现在60、50、38 d,之后种群数量开始剧增,建议在此期之前应采取防治措施。Abstract: Effects of environmental humidity on the growth, reproduction and survival of the Lasioderma serricorne population in laboratory were studied. Varied relative humidity, i. e., (50±5)%, (60±5)%, (70±5)%, (80±5)% and (90±5)% RH, at (32±1)℃ with a photoperiod of 14-h-L:10-h-D were applied. The results showed that (1) L. serricorne failed to completely develop a single generation under 50% or 90% RH-the first generation larvae died at 90% and the second instar perished at 50%, but it could under the other humidified conditions in the test; (2) there were no significant variations in the egg development time under the tested RHs, except 70% and 90%; (3) the larval developments were similar under 60% and 70% RH, however, at 80% RH, the duration of the 1st stage (i. e., 8.08 d), 2nd stage (i. e., 6.82 d), and 3rd stage (i. e., 7.05 d) were significantly shorter than the others; (4) the longest life span of 25.84 d was found in female adults at 60% RH, and 15.25 d in males at 80% RH; (5) the shortest pre-oviposition period of 2.55 d, the total pre-oviposition period of 58.41 d and the oviposition period of 7.00 d were found when the RH was 80%, but no significant difference at 60% or 70%; (6) there were significant differences on the fecundity, age-stage life expectancy (exj), and age-stage specific reproductive value (Vxj) of the insects reared under 60%, 70% or 80%, and they peaked at 60% RH; (7) the net reproduction rates (R0) decreased from 37.69 at 60% to 30.65 at 70% and 15.02 at 80% RH, whereas, the intrinsic rates of increase (r) and finite rates of increase (λ) showed no significant differences as the RH was altered; (8) there were no significant differences on the generation times (T) of 72.68 d at 60% and 72.79 d at 70% RH, but they were significantly longer than 62.80 d at 80%; and (9) the lowest larval counts were found on the 60th d under 60%, the 50th d under 70% and the 38th d under 80% RH, and escalated significantly afterward suggesting an optimal time for the insect control.
-
烟草甲Lasioderma serricorne (Fabricius)属鞘翅目Coleoptera、窃蠹科Anobiidae,是烟草仓储期的主要害虫, 在全世界广泛分布,国内绝大多数省区均有发生,特别是南方各省、市(自治区)发生较多[1-2]。其食性很杂,主要取食库藏烟叶,也为害各种粮食及其制品、茶叶、药材、干果、干肉、干鱼、皮毛、丝织品、纸张、动植物标本等[3-5]。烟草甲嗜食正在醇化的烟叶,烟草往往被蛀食成孔洞、残缺不全,严重时叶肉被蛀食光,仅剩叶脉,虫粪、丝网污染烟叶,严重影响烟草品级和商品价值[6],全国每年因该虫危害造成烟叶损失惨重,给烟草产业造成巨大的经济损失,成为世界烟草储藏的头号害虫[7-8]。
生命表的构建是研究种群存活和繁殖潜力的常用方法,通过生命表研究可较为准确地分析外界环境因子对昆虫种群动态、各虫态存活和发育、成虫繁殖力等的影响,并可对未来种群的发生进行预测,为农业害虫的防治提供指导,有力推动了害虫综合防控策略的发展[9-10]。根据昆虫的发生特点,不少学者建立了适应于昆虫种群研究的虫期组配生命表[11-13]。近年来,年龄-龄期两性生命表理论被广泛应用于昆虫生命表组建中[14-16]。相比于传统生命表,两性生命表将一特定昆虫种群的存活天数、死亡原因、龄期、产卵量等资料列表[17-19],得出特定种群-龄期的存活率、生殖力、净生殖力、净增殖率、内禀增长率、世代历期、生命期望值等相关指标,以此来分析引起该种群数量变动的环境因子,不仅能够精确地描述种群特征,而且还考虑到龄期分化和雄虫的影响[20-21]。
20世纪90年代以来,国内学者开展了烟草甲生物学特性、发生规律等相关研究[22],并基于传统生命表的方法开展了不同环境条件对烟草甲的影响,如王明洁等在4种温度(23.9、27.7、32.2、33.7℃)及3个湿度(RH 85%、76%、54%)组合条件下,分析了温湿度对烟草甲的影响[23];甘泳红等探讨了烟草甲试验种群的生物学参数并组建了烟草甲试验种群生命表[24]。研究表明烟草甲交配过的成虫明显比未交配过的成虫寿命长[25-26],因此在两性生命表组建实验中不论雌雄均需配对。低温和高温对烟草甲的生长发育影响较大[27-28]。本研究选择烟草甲的最适温度32℃作为温度条件,将烟草甲从卵到成虫分为8个发育龄期:卵期、幼虫期(1、2、3、4~6龄)、预蛹期、蛹期、雌雄成虫期,在5个不同相对湿度条件下通过组建和分析烟草甲两性生命表,旨在探讨不同湿度对烟草甲生长发育及繁殖的影响,以期对烟草仓库中烟草甲种群的预测和防治提供理论依据。
1. 材料与方法
1.1 材料
供试寄主:烟叶(品种为K36,购于福建省龙岩市烟草公司);供试虫源:烟草甲(来源于福建农林大学实验室饲养的实验种群);仪器设备:人工气候箱(宁波海曙赛福实验仪器公司,型号:PRX-25013), 体视显微镜(北京泰克仪器有限公司, 型号:XTL-20)等。
1.2 方法
试验条件:温度(32±1)℃,光周期14L:10D,光照度4 000 lx,5个不同湿度为:(50±5)%、(60±5)%、(70±5)%、(80±5)%和(90±5)%RH;试验方法:采用组建两性生命表的方法来研究不同湿度对烟草甲生长发育的影响,用烟叶对烟草甲饲养3代及以上,以确保烟草甲能在烟叶上完成1个世代。每种湿度条件各取同批次烟草甲24 h内所产的100粒卵作为生命表研究的起始虫量进行试验,每粒卵均置于垫有相同大小烟叶的培养皿(直径为6.0 cm, 皿盖上钻孔透气,皿盖和皿底之间用封口膜封死防止烟草甲逃逸),逐日观察记录烟草甲各个虫龄的死亡率及发育所需时间并更换烟叶。待成虫羽化后,雌虫优先与该组试验中同一时期羽化的雄虫配对,若无羽化雄虫则替补同条件下饲养的雄虫;若雄虫先于雌虫死亡则补雄虫,若雌虫先于雄虫死亡则不再记录雌虫存活天数和每日产卵量。雄虫需与该组实验中或同条件下饲养的雌虫进行配对,若雌虫先于雄虫死亡则补充雌虫,若雄虫先于雌虫死亡则不再记录雄虫存活天数。
1.3 数据统计分析与两性种群生命表的制作
数据利用Excel表进行初步统计,整理和分析;生命表参数和种群预测采用TWOSEX-MSChart程序分析[19];采用Bootstrap计算繁殖力和种群生命表参数的标准误等,并使用Sigmaplot 12.2软件进行作图;用SSPS 22.0进行单因素方差分析,采用LSD(最小差异显著法)比较各虫态(虫龄)的发育历期、寿命长短、产卵量等。将年龄-阶段两性种群生命表得出的参数通过TIMING-MSChart 2016程序来预测120 d后烟草甲的种群动态[29]。
由逐日记录的群体存活数和繁殖数来计算特定年龄阶段存活率(Sxj)即指一个新的个体存活到年龄x和龄期j的可能性;繁殖力fxj是指一头雌成虫在年龄x和龄期j所产的能够孵化的卵粒数。根据两性生命表理论,相关烟草甲实验种群两性生命表参数计算公式如下:
特定龄期存活率(lx):lx=m∑j=1sxj
特定年龄-龄期繁殖力(mx):mx=m∑j=1sxjfxjm∑j=1sxj
净增殖率(R0):R0=∞∑x=0lxmx
内禀增长率(r):∞∑x=0e−r(x+1)lxmx=1
世代历期(T):T=lnR0r
特定年龄-龄期生命期望值(exj):exj=∞∑i=x∞∑y=js′iy
繁殖力(vxj):υxj=er(x+1)sxj∞∑i=xe−r(i+1)m∑y=js′iyfiy
种群预测:ϕj,t=lg(nj,t+1+1)lg(nj,t+1)
rj,t=ln(nj,t+1+1nj,t+1)=ln(nj,t+1+1)−ln(nj,t+1) 2. 结果与分析
2.1 不同湿度对烟草甲种群存活率的影响
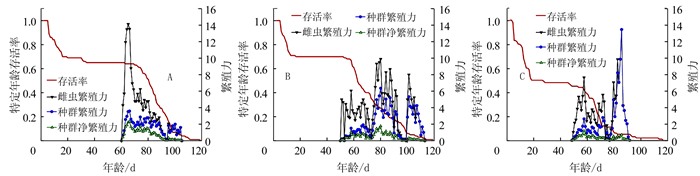
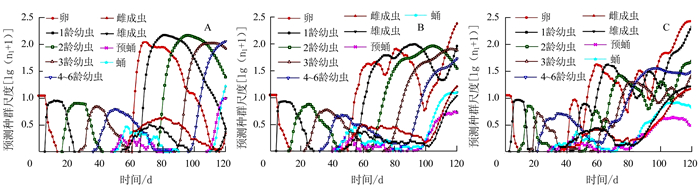
在5个湿度条件下烟草甲不同虫态的年龄-龄期存活率(Sxj)见图 1。图 1的曲线表明烟草甲从初产卵至能活到年龄x和龄期j的可能性。该曲线描绘出烟草甲卵、幼虫、蛹及成虫不同虫态的存活率以及不同龄期[卵、幼虫(1、2、3、4~6龄)、预蛹、蛹、雌雄成虫]间发育速率的差异。研究发现,卵期的孵化率(存活率)在80%RH条件下最高,为83%;90%RH条件下最低,为56%,其余则相差不大(图 1-A~E);在50%RH和90%RH条件下,烟草甲未能完成一个世代的发育,90%RH条件下幼虫在1龄全部死亡(图 1-E),50%RH条件下幼虫在2龄全部死亡(图 1-A),说明相对湿度高于90%或低于50%不利于烟草甲的生长发育。试验结果显示:烟草甲在60%RH、70%RH和80%RH条件下,新产的1粒卵成功发育为雌虫的几率分别为0.37、0.38和0.22,存活到雄成虫的几率分别为0.28、0.25和0.24;除卵期外,烟草甲在80%RH条件下各龄期的存活率均低于60%RH和70%RH条件下不同龄期的存活率。在80%条件下烟草甲卵期、幼虫期、预蛹期和蛹期的存活率分别为0.83、0.48、0.46和0.46;在60%RH下分别为0.81、0.65、0.65和0.65;在70%RH下分别为0.77、0.65、0.64和0.63。
![]() 图 1 不同相对湿度条件下烟草甲的特定年龄-龄期存活率(Sxj)注:A为50%RH,B为60%RH,C为70%RH,D为80%RH,E为90%RH。Figure 1. Age-stage specific survival rate (Sxj) of L. serricorne at varied RH
图 1 不同相对湿度条件下烟草甲的特定年龄-龄期存活率(Sxj)注:A为50%RH,B为60%RH,C为70%RH,D为80%RH,E为90%RH。Figure 1. Age-stage specific survival rate (Sxj) of L. serricorne at varied RH2.2 不同湿度对烟草甲发育历期的影响
不同湿度条件下,烟草甲整个发育阶段和不同虫态的持续时间有显著差异(表 1)。烟草甲卵的发育时间在70%RH条件下最短,为5.64 d,与60%RH(6.12 d)和80%RH(5.81 d)均无显著差异。1龄幼虫的发育历期在70%RH条件下最长为13.31 d,与60%RH(12.86 d)无显著差异,但在80%RH条件下发育历期最短为8.08 d,显著长于60%RH和70%RH下的同龄幼虫。2龄幼虫、3龄幼虫、蛹和成虫前期的发育时间在60%RH和70%RH条件下均无显著差异,但显著长于80%RH同虫龄虫态的发育历期;4~6龄幼虫的发育历期在70%RH和80%RH条件下差异显著,二者与60%RH无显著差异;预蛹的发育历期在60%RH和80%RH条件下无明显差异,但均显著长于70%RH的预蛹期。各虫态发育历期以预蛹期最短,其次是蛹期和卵期,幼虫期最长。60%RH和80%RH条件下4~6龄的发育历期比幼虫各个龄期都要长,可能是由于60%RH和80%RH条件下5龄和6龄幼虫偏多所致,且发育历期都较70%RH要长;70%RH在幼虫期各虫龄的发育历期无明显差异。
表 1 不同湿度下烟草甲卵、幼虫、预蛹和蛹的发育历期Table 1. Developmental durations of egg, larval and pupal stages of L. serricorne under varied RH龄期 发育历期/d 60%RH 70%RH 80%RH 卵 6.12±1.54ab(81) 5.64±1.06b(77) 5.81±0.92ab(83) 1龄幼虫 12.86±2.50a(69) 13.31±3.66a(70) 8.08±1.34b(66) 2龄幼虫 14.55±3.42a(65) 13.76±3.00a(70) 6.82±2.70b(50) 3龄幼虫 11.77±3.72a(65) 12.46±3.73a(65) 7.65±3.95b(49) 4~6龄幼虫 15.40±6.30ab(65) 12.58±8.37b(65) 18.96±8.88a(48) 预蛹 1.88±0.38a(65) 1.53±0.59b(64) 1.84±0.51a(46) 蛹 4.40±0.63a(65) 4.22±0.83a(63) 3.93±0.71b(46) 成虫前期 66.86±7.65a(65) 62.68±12.05a(63) 52.52±10.05b(46) 注:同行数据后不同小写字母表示经过LSD法检验差异显著(P<0.05);小括号内数字表示试虫存活数。表 2同。 2.3 不同湿度对烟草甲成虫寿命和繁殖力的影响
从表 2可以看出,烟草甲成虫寿命在70%RH和80%RH条件下差异不显著,但均显著短于60%RH下的成虫寿命。雌成虫寿命在70%RH和80%RH条件下无明显差异,均显著短于60%RH的雌成虫寿命;雄成虫寿命在70%RH和80%RH条件下无明显差异,但与60%RH下的成虫寿命差异显著。80%RH条件下雌成虫产卵前期最短为2.55 d,但均与60%RH和70%RH下的产卵前期无显著差异。产卵期在80%RH条件下最短为7.00d,均显著短于60%RH和70%RH下的产卵期。可育雌虫比率在60%RH和70%RH条件下无显著差异,但均显著低于80%RH下的可育雌虫比率。表 2数据分析还发现随着湿度的提高,60%RH、70%RH和80%RH等3个湿度条件下的产卵期、产卵量和性比均呈逐渐下降的趋势,60%RH和70%RH下雌虫的产卵期差异不显著,但二者均显著长于80%RH下的产卵期;60%RH条件下雌成虫平均产卵量显著多于70%RH和80%RH下的产卵量,三者的产卵量分别为110.88、92.85、68.32粒,每雌产卵量最高分别为224、253、124粒,每日单雌产卵量最高分别为57、63、53粒。3个湿度条件下成虫羽化后第1 d均不能产卵,产卵量从第2 d开始逐渐增加,产卵主要集中在羽化后的第4~10 d,之后产卵量很小,产卵量最高主要在第5~7 d。结合成虫寿命和产卵前期长短,可以看出在适宜温度范围内湿度越低越有利于烟草甲成虫的存活和产卵。
表 2 不同湿度下烟草甲的成虫寿命和雌虫繁殖力Table 2. Adult lifespan and female fecundity of L. serricorne under varied RH参数 相对湿度 60%RH 70%RH 80%RH 成虫寿命/d 21.42±7.77a(65) 16.81±8.07b(63) 18.48±7.61ab(46) 雌成虫寿命/d 25.84±7.03a(37) 19.29±8.66b(38) 20.91±9.46b(22) 雄成虫寿命/d 15.57±3.86b(28) 13.04±5.27a(25) 16.25±4.56a(24) 产卵前期/d 3.06±0.90a(33) 3.15±0.97a(33) 2.55±1.92a(22) 产卵期/d 9.27±2.14a(33) 8.55±4.40a(33) 7.00±3.28b(22) 产卵量/粒(雌) 110.88±55.68a(33) 92.85±73.93ab(33) 68.32±40.81b(22) 性比(雄:雌) 0.78±0.20b 0.68±0.18b 1.14±0.36a 可育雌虫比率(产雌:总雌) 0.89±0.051b 0.87±0.06b 1.00±0.00a 繁殖力曲线由曲线mx、lxmx、fx8共同组成(图 2)。其中lx是指不考虑龄期分化的种群年龄-特征存活率;特定年龄-阶段繁殖力mx是指烟草甲种群在年龄x时的平均产卵量;种群特定年龄-阶段特征净繁殖力lxmx为种群年龄特征存活率lx与种群年龄-阶段特征繁殖力mx的乘积。fx8表示雌成虫在年龄x时的所产的可孵化卵粒数。60%RH、70%RH和80%RH等3个湿度条件下的lx曲线具备相同的趋势,即前期曲线较陡,后期趋于平缓,表明不同湿度下烟草甲种群内个体死亡主要发生在整个发育阶段的前面阶段,使得存活率lx在种群发育的前期变化较大,而后期种群内个体的死亡情况趋于缓和,试验结果也印证了这个结论:即3个湿度条件下烟草甲种群中1龄幼虫死亡率均为最高(80%RH下2龄幼虫也较高),之后种群死亡率极低,表明烟草甲幼虫只要能活过2龄即多可成功化蛹并羽化为成虫繁殖下一代。繁殖力曲线(mx、lxmx、fx8)最早出现在80%RH,为第49 d,70%RH为第50 d,60%RH最晚出现,为第60 d。特定年龄生殖力mx随着湿度的升高而减少,但特定年龄净繁殖力lxmx随湿度的升高而增加,3个湿度条件下的产卵生殖期分别为42、62、44 d。60%RH的fx8曲线前期很陡,而70%RH和80%RH的曲线则前期相对平缓,说明60%RH雌成虫羽化和产卵均较为集中,而70%RH和80%RH下雌成虫羽化和产卵均不集中,导致曲线出现忽高忽低的情形,3个湿度条件下雌虫特定年龄繁殖力的峰值分别出现在64、79、85 d;70%RH对应的烟草甲产卵高峰期最长,60%RH最短。
![]()
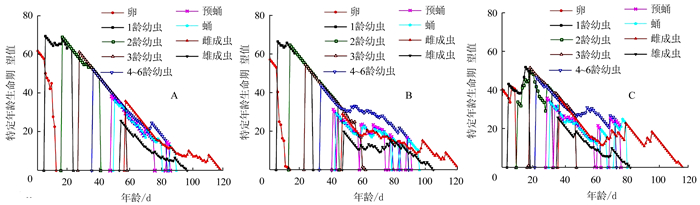
2.4 不同湿度对烟草甲的生命期望值的影响
特定年龄-龄期生命期望值(exj)表示对于x年龄j龄期的个体在未来的期望存活时间(图 3)。60%RH条件下的每个龄期阶段的起始生命期望值exj均显著长于70%RH和80%RH,说明烟草甲在60%RH条件下生长发育相对缓慢些。70%RH的卵期到3龄的起始生命期望值要高于80%RH,而4~6龄到雄成虫的起始生命期望值要低于80%RH。3个湿度条件下卵的起始生命期望值分别为61.79、57.01、40.17 d(图 3-A~C)。
![]() 图 3 不同相对湿度条件下烟草甲特定年龄-龄期生命期望值(exj)Figure 3. Age-stage specific life expectancy (exj) of L. serricorne under varied RH
图 3 不同相对湿度条件下烟草甲特定年龄-龄期生命期望值(exj)Figure 3. Age-stage specific life expectancy (exj) of L. serricorne under varied RH2.5 不同湿度对烟草甲繁殖率贡献值的影响
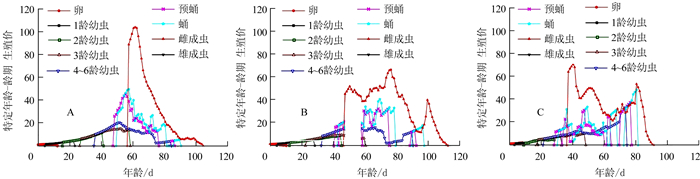
特定年龄-龄期生殖价(Vxj)是描述x年龄j龄期的个体对将来种群增长的贡献率(图 4)。结果显示, 随着湿度的增加,烟草甲卵期到蛹期的生殖价均增长越慢。60%RH、70%RH和80%RH等3个湿度条件下, 第一个雌成虫生殖价Vxj值分别为89.12、45.38、64.25,分别出现在57、47、37 d。随着湿度的增加,雌成虫的生殖价最高峰即最大贡献率分别出现在61、76、40 d,对应的生殖价Vxj[V(61, 8)、V(76, 8)、V(40, 8)]值分别为104.32、66.76、71.01。70%RH和80%RH条件下雌成虫均出现多个生殖价高峰,3个湿度条件下雌成虫的生殖价持续时间分别为47、65、54 d。
![]() 图 4 不同相对湿度条件下烟草甲特定年龄-龄期生殖价(vxj)Figure 4. Age-stage specific reproductive value (vxj) of L. serricorne under varied RH
图 4 不同相对湿度条件下烟草甲特定年龄-龄期生殖价(vxj)Figure 4. Age-stage specific reproductive value (vxj) of L. serricorne under varied RH2.6 不同湿度条件下烟草甲的种群参数
烟草甲在60%RH、70%RH和80%RH条件下的实验种群参数:净增殖率(R0)、内禀增长率(r)、周限增长率(λ)、平均世代历期(T)的平均值和标准误见表 3。结果表明湿度对净增殖率(R0)和世代周期(T)影响显著。净增殖率(R0)在60%RH和70%RH条件下无显著差异,但显著高于80%RH(最低值为15.02);3个湿度对应的内禀增长率(r)和周限增长率(λ)随着湿度的升高略呈下降趋势,但并无显著差异;虽然80%RH条件下种群净增殖率(R0)最低,但世代周期(T)最短为62.80 d,显著短于60%RH和70%RH条件下的世代周期。因此随着时间的延续,烟草甲种群依然可以不断地快速扩增,即烟草甲在60%RH、70%RH和80%RH条件下,若其他环境因子适合,种群将分别以0.05、0.05和0.04的指数增长速率在1个世代周期为72.68、72.79、62.8 d内每天约增长1.05倍。
表 3 不同湿度下烟草甲的种群参数Table 3. Population parameters of L. serricorne under varied RH种群参数 相对湿度 60%RH 70%RH 80%RH 净增殖率(R0) 37.69±6.15a 30.65±6.06a 15.02±3.39b 内禀增长率(r,d-1) 0.05±0.00a 0.05±0.00a 0.04±0.00a 周限增长率(λ,d-1) 1.05 ±0.003a 1.05±0.003a 1.04±0.004a 世代周期(T,d-1) 72.68±1.47a 72.79±2.33a 62.80±3.11b 2.7 种群预测
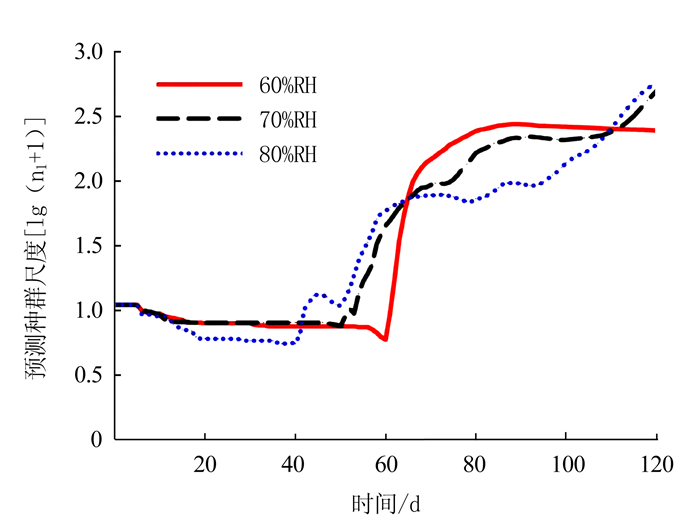
种群增长趋势预测结果可为害虫的综合治理提供理论依据。本研究基于两性生命表以100粒卵为起始虫数,模拟预测了未来120 d(4个月)内烟草甲种群增长的情况。结果表明,60%RH、70%RH和80%RH等3个湿度下烟草甲未来种群最低虫口数分别出现在60、50、38 d,之后种群数量开始急剧增加,并随着湿度的增加越来越快(图 5),建议在种群数量增加之前对烟草甲采取适当的防治措施。烟草甲在3个不同湿度条件下取食烟叶,至120 d时种群数量明显不同,60%RH下不足2.5个数量单位,而70%RH和80%RH下则超过了2.5个数量单位,其中80%RH下其种群数量最高达到了2.8个数量单位。每隔一段时间60%RH和80%RH的种群预测曲线交替增长,70%RH则处在两者之间,在40、65、110 d,3个湿度条件的种群数量曲线便交汇在一起,其种群数量大致相同,表明60%RH-80%RH均为烟草甲的最适湿度(图 5)。4个月后,60%RH下烟草甲种群中刚刚出现第2代成虫,而70%RH和80%RH下种群已处于第3代,且具有世代重叠现象(图 6)。
![]() 图 5 不同相对湿度条件下烟草甲的种群预测(总种群尺度)Figure 5. Population projection (total population size) of L. serricorne reared on tobacco at varied RH
图 5 不同相对湿度条件下烟草甲的种群预测(总种群尺度)Figure 5. Population projection (total population size) of L. serricorne reared on tobacco at varied RH![]() 图 6 不同相对湿度条件下烟草甲的种群预测(各龄期种群尺度)Figure 6. Population projection (stage-wise population size) of L.serricorne reared on tobacco at varied RH
图 6 不同相对湿度条件下烟草甲的种群预测(各龄期种群尺度)Figure 6. Population projection (stage-wise population size) of L.serricorne reared on tobacco at varied RH3. 讨论与结论
烟草甲是烟叶仓储期最重要的害虫,温度和湿度均是影响烟草甲种群发展的重要因素[23]。本研究在最适温度32℃条件下,通过构建实验种群两性生命表的方法探讨不同湿度对烟草甲生长发育和繁殖的影响,结果表明,50%RH和90%RH条件下烟草甲均未能完成一代的发育,湿度低于50%或高于90%不利烟草甲的生长发育,考虑到湿度过大对烟草存储不利,建议为控制烟草甲对烟叶的为害,烟仓内的相对湿度宜调到50%RH以下。本研究结果表明,60%~80%RH为烟草甲适宜的湿度范围,这与张孝羲等得出的最适湿度在70%~85%基本吻合[4],之间的差距可能是由于所使用人工气候箱湿度的正负误差、寄主条件和光周期不同所导致。此外本试验中烟草甲卵期除70%RH和90%RH差异显著,其余湿度均无明显差异,这也与前人的研究结果大体一致。本研究发现3个湿度条件下,1~3龄幼虫各阶段发育时间相近,但80%RH条件下1~3龄各龄幼虫发育时间比60%RH和70%RH的均显著缩短。前人研究表明,随着虫龄的增长,4~6龄各个虫龄的发育历期会越来越短[4-5],这与本试验所观察到的结果相一致。研究结果表明,3个湿度条件下烟草甲种群中幼虫以4龄为主,6龄占比较少,7龄未出现,这与冯小明等的烟草甲幼虫多为5、6龄,少数出现4、7龄的结果不相吻合[5]。本研究发现烟草甲预蛹期非常短,3个湿度条件下均不到2.0 d,这与张孝羲等得出的预蛹期为约7 d[4]差异明显,原因尚待进一步分析。甘泳红等研究表明蛹期时间约为3.61 d[24],这与本研究80%RH下最短的蛹期(3.93 d)较为接近。有资料表明雌成虫羽化后3~5 d才能交配产卵[5],但本研究却发现羽化后2 d烟草甲即可交配产卵。本研究结果表明, 烟草甲雄成虫可存活10~20 d,但有少数雄成虫能超过20 d,在80%RH条件下雄成虫寿命最长可达到28 d;关于雌成虫的寿命,本研究结果与前人的基本一致,即烟草甲雌成虫可存活20~40 d且80%卵产于第5~10 d,雌成虫寿命一般高于雄成虫[30]。冯小明等表明烟草甲雌成虫产卵范围约为50~95粒[5], 与本试验70%RH和80%RH条件下的产卵量基本一致,但在60%RH条件下,每雌平均产卵量最大,为110.88粒;以往研究表明烟草甲产卵最多不超过100粒,但本试验3个湿度条件下最大产卵量均超过100粒,且在60%和70%RH条件下烟草甲最大产卵量已经超过200粒。
有报道认为,湿度对内禀增长率r、净增殖率R0、平均世代周期T影响显著[23],这与本试验结果不完全一致。本研究发现,3个湿度条件下烟草甲的内禀增长率r和周限增长率λ无明显差异,而80%RH下的净增殖率R0、平均世代周期T显著低于(短于)60%RH和70%RH下,表明湿度会显著影响烟草甲种群的增殖和世代周期的长短。通过比对存活率、繁殖力、生殖价、净增殖率等数据可以发现,60%RH条件下烟草甲种群生长发育和繁殖均处于最优状态,理论上60%RH条件下烟草甲拥有较高的存活率、预期寿命更长、较大的生育价值和较高的净增殖率能让种群更好的扩展。但种群预测结果发现:相较于80%RH,60%RH条件下烟草甲的世代周期时间过长,而80%RH下较短的世代周期却也能让种群数量快速增殖,3个湿度条件在一定时间后种群数量会趋于相同,并且最终80%RH和70%RH下的烟草甲种群数量将反超过60%RH,表明60%、70%、80%RH均适于烟草甲生长发育和繁殖。
对仓储害虫烟草甲而言,湿度对其种群的生存和繁衍具有重要影响。本试验研究了不同湿度50%、60%、70%、80%、90%RH下烟草甲的存活率、发育历期和种群生命表参数,对烟草甲种群动态预测具有重要意义。除湿度外,温度也是影响自然仓储生态环境下烟草甲种群动态的一个关键因素[23],同时其他生物因素和非生物因素也均影响着烟草甲的种群变化。因此没有外界干扰(天敌)的室内可控条件下得出的烟草甲种群动态与实际烟草仓库中的种群动态有一定的差异,今后可进一步模拟真正烟仓的生态环境开展相关的研究,为深入了解烟仓或其他仓库中烟草甲的发生规律及综合防治提供理论依据。
-
![]()
图 1 不同相对湿度条件下烟草甲的特定年龄-龄期存活率(Sxj)
注:A为50%RH,B为60%RH,C为70%RH,D为80%RH,E为90%RH。
Figure 1. Age-stage specific survival rate (Sxj) of L. serricorne at varied RH
![]()
![]()
图 3 不同相对湿度条件下烟草甲特定年龄-龄期生命期望值(exj)
Figure 3. Age-stage specific life expectancy (exj) of L. serricorne under varied RH
![]()
图 4 不同相对湿度条件下烟草甲特定年龄-龄期生殖价(vxj)
Figure 4. Age-stage specific reproductive value (vxj) of L. serricorne under varied RH
![]()
图 5 不同相对湿度条件下烟草甲的种群预测(总种群尺度)
Figure 5. Population projection (total population size) of L. serricorne reared on tobacco at varied RH
![]()
图 6 不同相对湿度条件下烟草甲的种群预测(各龄期种群尺度)
Figure 6. Population projection (stage-wise population size) of L.serricorne reared on tobacco at varied RH
表 1 不同湿度下烟草甲卵、幼虫、预蛹和蛹的发育历期
Table 1 Developmental durations of egg, larval and pupal stages of L. serricorne under varied RH
龄期 发育历期/d 60%RH 70%RH 80%RH 卵 6.12±1.54ab(81) 5.64±1.06b(77) 5.81±0.92ab(83) 1龄幼虫 12.86±2.50a(69) 13.31±3.66a(70) 8.08±1.34b(66) 2龄幼虫 14.55±3.42a(65) 13.76±3.00a(70) 6.82±2.70b(50) 3龄幼虫 11.77±3.72a(65) 12.46±3.73a(65) 7.65±3.95b(49) 4~6龄幼虫 15.40±6.30ab(65) 12.58±8.37b(65) 18.96±8.88a(48) 预蛹 1.88±0.38a(65) 1.53±0.59b(64) 1.84±0.51a(46) 蛹 4.40±0.63a(65) 4.22±0.83a(63) 3.93±0.71b(46) 成虫前期 66.86±7.65a(65) 62.68±12.05a(63) 52.52±10.05b(46) 注:同行数据后不同小写字母表示经过LSD法检验差异显著(P<0.05);小括号内数字表示试虫存活数。表 2同。  下载: 导出CSV
下载: 导出CSV
表 2 不同湿度下烟草甲的成虫寿命和雌虫繁殖力
Table 2 Adult lifespan and female fecundity of L. serricorne under varied RH
参数 相对湿度 60%RH 70%RH 80%RH 成虫寿命/d 21.42±7.77a(65) 16.81±8.07b(63) 18.48±7.61ab(46) 雌成虫寿命/d 25.84±7.03a(37) 19.29±8.66b(38) 20.91±9.46b(22) 雄成虫寿命/d 15.57±3.86b(28) 13.04±5.27a(25) 16.25±4.56a(24) 产卵前期/d 3.06±0.90a(33) 3.15±0.97a(33) 2.55±1.92a(22) 产卵期/d 9.27±2.14a(33) 8.55±4.40a(33) 7.00±3.28b(22) 产卵量/粒(雌) 110.88±55.68a(33) 92.85±73.93ab(33) 68.32±40.81b(22) 性比(雄:雌) 0.78±0.20b 0.68±0.18b 1.14±0.36a 可育雌虫比率(产雌:总雌) 0.89±0.051b 0.87±0.06b 1.00±0.00a
下载: 导出CSV
表 3 不同湿度下烟草甲的种群参数
Table 3 Population parameters of L. serricorne under varied RH
种群参数 相对湿度 60%RH 70%RH 80%RH 净增殖率(R0) 37.69±6.15a 30.65±6.06a 15.02±3.39b 内禀增长率(r,d-1) 0.05±0.00a 0.05±0.00a 0.04±0.00a 周限增长率(λ,d-1) 1.05 ±0.003a 1.05±0.003a 1.04±0.004a 世代周期(T,d-1) 72.68±1.47a 72.79±2.33a 62.80±3.11b
下载: 导出CSV
-
[1] RYAN L. Post-harvest tobacco infestation control[M]. London:Chapman & Hall, 1995:155.
[2] 白旭光.储藏物害虫与防治[M].北京:科学出版社, 2002:294-295. [3] 柳琼友, 顾丁, 陈文龙.烟草甲的防治研究进展[J].湖北农业科学, 2007, (5):841-844. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=hbnykx200705065 [4] 张孝羲, 王明洁.烟草甲的实验生态研究[J].昆虫学报, 1996, 39(4):383-392. http://www.cnki.com.cn/Article/CJFDTOTAL-KCXB604.007.htm [5] 冯小明, 王方晓.烟草甲的生物学特性研究[J].昆虫知识, 1997, 34(2):85-87. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=zbjsytg200809003 [6] SUN X, WANG L F, FENG Y, et al. A case report:A rare case of infant gastrointestinal canthariasis caused by larvae of Lasioderma serricorne (Fabricius, 1792) (Coleoptera:Anobiidae)[J]. Infectious Diseases of Poverty, 2016, (5):34. DOI: 10.1186%2Fs40249-016-0129-6
[7] 罗梅浩, 马淑健, 殷花娥, 等.烟草甲Lasioderma serricorne Fabricius对储存烟叶为害损失的初步研究[J].河南农业大学学报, 2003, 37(3):249-252. http://www.cqvip.com/Main/Detail.aspx?id=8415308 [8] AKINKUROLERE R O, 张宏宇, 饶琼, 等.烟草甲在三种寄主上存活、发育的研究[J].粮食储藏, 2006, 34(5):8-10, 14. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=lscc200605002 [9] GABRE R A, ADHAM F K, CHI H. Life table of Chrysomya megacephala (Fabricius) (Diptera:Calliphoridae)[J]. Acta Oecologica, 2005, (27):179-183. https://www.cambridge.org/core/journals/bulletin-of-entomological-research/article/div-classtitlefactors-influencing-oviposition-larval-growth-and-mortality-in-span-classitalicchrysomya-megacephalaspan-diptera-calliphoridae-a-pest-of-salted-dried-fish-in-south-east-asiadiv/5CF8BE0602C2B9A7A0EF16D85A493EF0
[10] 吴坤君, 龚佩瑜, 李秀珍, 等.粘虫的年龄-发育期结构生命表[J].昆虫学报, 1994, 37(4):425-434. http://cdmd.cnki.com.cn/Article/CDMD-10422-2008189247.htm [11] 王海鸥, 薛瑶, 雷仲仁.恒温和波动温度下西花蓟马的实验种群生命表[J].中国农业科学, 2014, 47(1):61-68. http://www.chinaagrisci.com/article/2014/0578-1752-47-1-61.html [12] 庞雄飞, 侯任环, 包华理.褐飞虱子种群生命表的组建方法[J].华南农业大学学报, 1992, 13(1):1-5. http://www.oalib.com/paper/4843979 [13] 高增祥, 徐汝梅, 吴子江, 等.基于生命表参数的棉铃虫种群动态模拟研究[J].昆虫学报, 2005, 48(4):568-575. http://www.cqvip.com/qk/94065X/200504/18057012.html [14] HUANG Y B, CHI H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera:Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations[J]. Insect Science, 2012(19):263-273.
[15] ZHENG X M, TAO Y L, Chi H, et al. Adaptability of small brown planthopper to four rice cultivars using life table and population projection method[J]. Scientific Reports, 2017, (7):42399;doi: 10.1038/srep42399.
[16] YANG Y T, LI W X, XIE W, et al. Development of Bradysia odoriphaga (Diptera:Sciaridae) as affected by humidity:an age-stage, two-sex, life-table study[J]. Applied Entomology and Zoology, 2015, 50:3-10. DOI: 10.1007/s13355-014-0295-6
[17] PENG L, MIAO Y X, HOU Y M. Demographic comparison and population projection of Rhynchophorus ferrugineus (Coleopt-era:Curculionidae) reared on sugarcane at different temperatures[J]. Scientific Reports, 2016, (6):31659; oi:10.1038/srep31659. http://europepmc.org/articles/PMC4992881/
[18] PAVEL S. Prey With Glyphosate Does Not Alter the Demographic Parameters and Predation of the Harmonia axyridis (Coleoptera:Coccinellidae)[J]. Journal of Economic Entomology, 2017:1-8. http://www.chinaagrisci.com/Jwk_zgnykxen/EN/rss_zxly.xml
[19] CHI H. Life-table analysis incorporating both sexes and variable evelopment rates among individuals[J]. Environmental Entomology, 1988, (17):26-34. http://cn.bing.com/academic/profile?id=7a4427517e58535ef9d5949b7b2369df&encoded=0&v=paper_preview&mkt=zh-cn
[20] CHI H, GETZ W M. Mass rearing and harvesting based on an age-stage, two-sex life table:a potato tuber-worm (Lepidoptera:Gelechiidae) case study[J]. Environmental Entomology, 1988, (17):18-25. https://www.researchgate.net/publication/233707464_Mass_Rearing_and_Harvesting_Based_on_an_Age-Stage_Two-Sex_Life_Table_A_Potato_Tuberworm_Lepidoptera_Gelechiidae_Case_Study
[21] CHI H, LIU H. Two new methods for the study of insect population ecology[J]. Bulletin of the Institute of Zoology Academia Sinica Monograph, 1985, (24):225-240. http://cn.bing.com/academic/profile?id=d3b54f8531bc23779b322fe29dc2d196&encoded=0&v=paper_preview&mkt=zh-cn
[22] 吕建华, 袁良月.烟草甲生物学特性研究进展[J].中国植保导刊, 2008, (9):12-15. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=kjzxdb200925007 [23] 王明洁, 张孝羲.烟草甲实验种群特定时间生命表的研究[J].南京农业大学学报, 1995, (4):52-56. http://www.cqvip.com/QK/85595X/199602/3001131587.html [24] 甘泳红, 刘光华, 周长青, 等.烟草甲试验种群生物学参数及生命表研究[J].广东农业科学, 2011, (11):86-89. DOI: 10.3969/j.issn.1004-874X.2011.11.033 [25] 李景科, 高梅香, 张雪萍.昆虫的寿命研究[J].哈尔滨师范大学:自然科学学报, 2005, 21(6):79-86. http://mall.cnki.net/magazine/Article/HEBY200506024.htm [26] MAHROOF R M, PHILLIPS T W. Life history parameters of Lasiodermo serricome (F.) as influenced by food sources[J]. Journal of Stored Products Research, 2008, 44:219-226. DOI: 10.1016/j.jspr.2007.12.001
[27] 王秀芳, 任广伟, 周显升, 等.低温对不同虫态烟草甲的影响[J].华北农学报, 2010, 25(S1):287-289. DOI: 10.7668/hbnxb.2010.S1.065 [28] 吕建华, 钟建军, 张会娜, 等.高温处理对烟草甲各虫态的致死作用研究[J].农业灾害研究, 2014, 4(3):15-17. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=nzyj201403008&dbname=CJFD&dbcode=CJFQ [29] CHI H. TWOSEX-MSChart: A computer program for the life tables and population growth of insect[OL]. National Chung Hsing University, Taichung, Taiwan. 2015. http://140.120.197.173/Ecology/.
[30] 王方晓, 魏重生, 冯小明.烟草甲的研究[J].中国烟草科学, 1998, (2):47-49. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGYV802.014.htm -
期刊类型引用(1)
1. 朱婷,马玲,冯灏怡,邓楠,刘心亮,刘明骞. 离子型稀土矿废弃地剑豆耐性种质筛选. 热带亚热带植物学报. 2024(04): 465-474 .  百度学术
百度学术
其他类型引用(2)
计量
- 文章访问数: 1871
- HTML全文浏览量: 333
- PDF下载量: 13
- 被引次数: 3
