

Formation of Proteus mirabilis Persister in Chicken under Gentamicin Sulphate Treatment
-
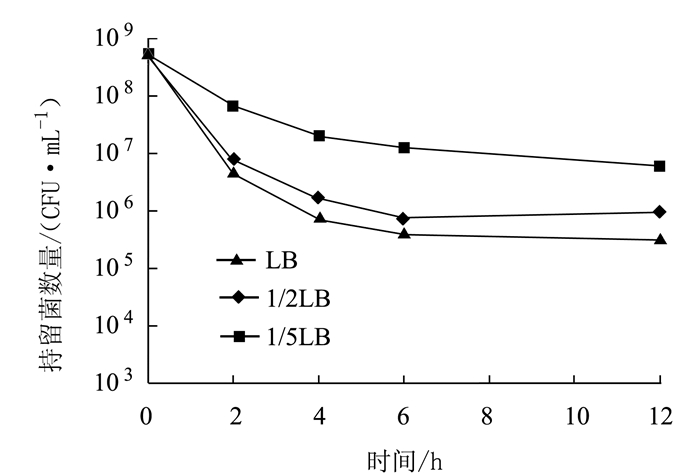
摘要: 为了解奇异变形杆菌持留菌形成特征和防控家禽养殖中细菌性疾病,以鸡源致病菌奇异变形杆菌PM2658为研究对象,通过二倍稀释法测得抗生素对鸡源奇异变形杆菌PM2658的最小抑菌浓度(Minimum inhibitory of concentration,MIC),用斑点培养(spot-plate)法分析PM2658在不同抗生素和营养条件下持留菌形成特征。结果显示,硫酸庆大霉素、头孢噻肟钠、氧氟沙星对PM2658的MIC值分别为1.56、0.78和3.13 μg·mL-1。在16×MIC和100×MIC硫酸庆大霉素处理下,鸡源奇异变形杆菌PM2658都可以形成持留菌,形成率分别为8.04×10-4和1.03×10-6。16×MIC硫酸庆大霉素-4×MIC头孢噻肟钠、16×MIC硫酸庆大霉素-4×MIC氧氟沙星、4×MIC头孢噻肟钠-4×MIC氧氟沙星、16×MIC硫酸庆大霉素-4×MIC头孢噻肟钠-4×MIC氧氟沙星不同抗生素组合处理下,鸡源奇异变形杆菌PM2658的持留菌形成规律与单独硫酸庆大霉素处理的持留菌形成规律一致,也表明鸡源奇异变形杆菌PM2658持留菌具有多重抗生素耐受性。在16×MIC硫酸庆大霉素处理下,与LB和1/2 LB培养条件相比,1/5 LB培养条件下鸡源奇异变形杆菌持留菌形成率最高,比前两者分别提高了17.67倍和5.05倍,说明鸡源奇异变形杆菌持留菌的形成还与营养环境密切相关。持留菌遗传学特性检测结果显示,鸡源奇异变形杆菌PM2658持留菌的遗传学特性并没有发生改变。鸡源奇异变形杆菌确实可以形成持留菌,其形成特征取决于特定的抗生素条件。Abstract: For understanding the biological characteristics of persister formation of Proteus mirabilis, and control and prevention of bacterial diseases in poultry, Proteus mirabilis PM2658 from chicken was used as materials. Minimum inhibitory of concentrations (MIC) of antibiotics to Proteus mirabilis PM2658 were determined by two-fold dilution method. Under different antibiotic treatments and nutritional conditions, the characteristics of persister formation of Proteus mirabilis were determined by spot-plate protocol. Results showed that MIC of gentamicin sulphate (Gen), cefotaxime sodium (Cef) and ofloxacin (Ofx) to Proteus mirabilis PM2658 were 1.56, 0.78 and 3.13 μg·mL-1, respectively. Treated by 16×MIC and 100×MIC of Gen, stationary phase of Proteus mirabilis PM2658 could form persister, and the percentages of persister formation were 8.04×10-4 and 1.03×10-6, respectively. Moreover, the regularity of peresister formation under antibiotic combinations of 16×MIC Gen-4×MIC Cef, 16×MIC Gen-4×MIC Ofx, 4×MIC Cef-4×MIC Ofx, 16×MIC Gen-4×MIC Cef-4×MIC Ofx was the same as that of single Gen treatment, indicating that persister of Proteus mirabilis PM2658 had multiple drug tolerance. The rate of formation of Proteus mirabilis PM2658 persister in 1/5 LB was higher than those in LB and 1/2 LB under 16×MIC Gen treatment, which separately increased by 17.67 times and 5.05 times, which indicated that the formation of Proteus mirabils PM2658 persister was related to nutrition of environment. The heritage of Proteus mirabilis PM2658 persister did not change, which was further determined after every different treatments mentioned above. Taken together, Proteus mirabilis from chicken do form persister, and characteristics of persister formation depends on specific antibiotic.
-
Keywords:
- Proteus mirabilis /
- drug tolerance /
- persister /
- gentamicin sulphate
-
随着养兔业的规模化发展,呼吸道疾病已成为制约其发展的重要因素之一。其中,由金黄色葡萄球菌和支气管败血波氏杆菌感染导致的呼吸道疾病尤为突出。部分兔场由金黄色葡萄球菌感染导致的发病率约30%,病兔死亡率达90%以上[1];而支气管败血波氏杆菌的感染率可高达70%,导致感染兔生产性能低下[2]。由金黄色葡萄球菌和支气管败血波氏杆菌感染导致的呼吸道疾病具有相似的临床症状,如喷嚏、咳嗽、流鼻涕、严重者流脓性鼻涕;剖解均可见气管出血、肺脏出血坏死,严重者则表现为化脓性肺炎[3-6]。此外,在多数病例中还可见这两种病原菌的混合感染,给临床鉴别诊断带来困难[6]。
目前,针对兔金黄色葡萄球菌和支气管败血波氏杆菌的检测方法主要有细菌分离鉴定、血清学检测和单重PCR等[6-8],还未见能同时检测这两种兔源病原菌的双重PCR检测方法。本试验根据金黄色葡萄球菌的nuc基因和支气管败血波氏杆菌的fimN基因保守序列,设计2对特异性引物,建立兔金黄色葡萄球菌和支气管败血波氏杆菌双重PCR检测方法,以期为两种兔病原菌的鉴别诊断提供技术支持。
1. 材料与方法
1.1 试验材料
1.1.1 菌株
兔金黄色葡萄球菌、支气管败血波氏杆菌、多杀性巴氏杆菌、大肠杆菌和魏氏梭菌均由本实验室分离保存。
1.1.2 主要试剂
DL2000 Marker和胶回收试剂盒购自宝生物工程(大连)有限公司;2 × Easy Taq PCR SuperMix和细菌基因组DNA提取试剂盒购自北京全式金生物技术有限公司。
1.2 试验方法
1.2.1 引物设计与合成
根据GenBank中已发表的金黄色葡萄球菌nuc基因序列和支气管败血波氏杆菌fimN基因序列,利用Primer Premier 5.0软件设计2对特异性引物。引物由上海博尚生物技术有限公司合成,引物序列和扩增片段长度见表 1。
表 1 双重PCR引物Table 1. Primers used for duplex PCR assay病原 靶基因 引物序列(5′-3′) 参考序列登录号 产物大小/bp 金黄色葡萄球菌 nuc F:ttggttgatacacctgaaacaa;R:cgttgtcttcgctccaaata DQ399678 322 支气管败血波氏杆菌 fimN F:gtgatcaccggcaacatc;R:tgcctgtggctttcttca AF231910 478 1.2.2 单重PCR扩增与测序鉴定
以金黄色葡萄球菌和支气管败血波氏杆菌基因组DNA为模板,利用单引物进行PCR扩增。PCR扩增体系:2 × PCR Mix 25 μL,细菌基因组DNA 2 μL,上下游引物各2 μL,ddH2O补齐至50 μL。PCR反应程序:94℃预变性4 min;94℃ 30 s、56℃ 30 s、72℃ 30 s,30个循环;72℃延伸10 min。目的片段经1.5%琼脂糖电泳回收并送上海博尚生物技术有限公司测序。
1.2.3 双重PCR反应体系的建立及优化
将金黄色葡萄球菌和支气管败血波氏杆菌基因组DNA的质量浓度均调整至50 ng·μL-1,等体积混合,作为双重PCR的模板。将nuc基因上下游引物和fimN基因上下游引物均稀释至20 μmol·L-1,等体积混合作为双重PCR的引物。PCR扩增体系:2 × PCR Mix 25 μL,混合基因组DNA 2 μL,混合引物4 μL,ddH2O补齐至50 μL。PCR反应程序:94℃预变性4 min;94℃ 30 s、56℃ 30 s、72℃ 30 s,30个循环;72℃延伸10 min。在此基础上,将退火温度设置为53~60℃,混合引物终浓度设置为0.2、0.3、0.4、0.5、0.6 μmol·L-1对双重PCR反应条件进行优化,确定最佳的退火温度和引物浓度。
1.2.4 双重PCR的特异性试验
应用建立的双重PCR方法,对兔金黄色葡萄球菌、支气管败血波氏杆菌、多杀性巴氏杆菌、大肠杆菌和魏氏梭菌的基因组DNA进行PCR扩增,同时设置阴性对照,验证双重PCR检测方法的特异性。
1.2.5 双重PCR的敏感性试验
将金黄色葡萄球菌和支气管败血波氏杆菌基因组DNA的浓度均调整至50 ng·μL-1,等体积混合后进行10倍倍比稀释,以稀释的混合基因组DNA为模板,应用建立的双重PCR方法进行PCR扩增,检验该方法的敏感性。
1.2.6 临床样品的检测
63份已知结果的分离株或分离株混合样品,其中20份为金黄色葡萄球菌阳性,24份为支气管败血波氏杆菌阳性,14份为金黄色葡萄球菌和支气管败血波氏杆菌混合感染,5份为金黄色葡萄球菌和支气管败血波氏杆菌阴性样品。应用建立的双重PCR方法检测上述63份临床样品,比较该方法与已知结果的一致性。
2. 结果与分析
2.1 单重PCR扩增与测序鉴定
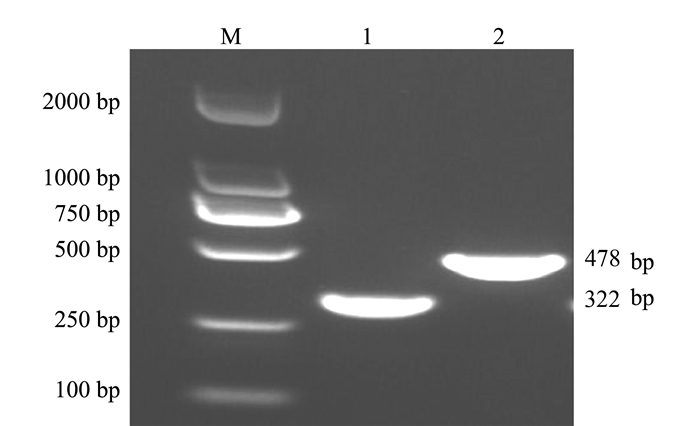
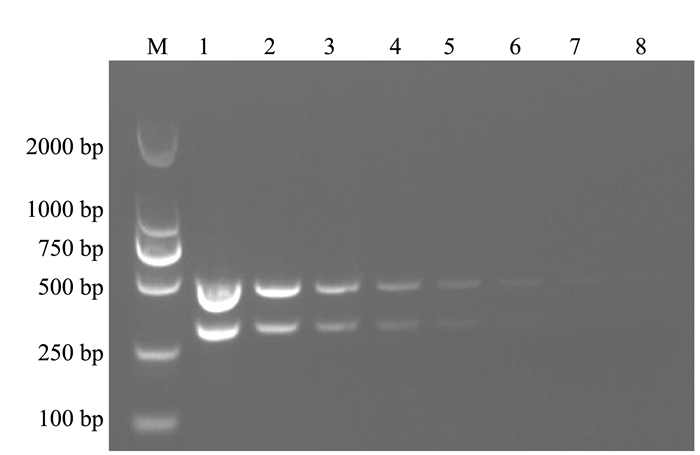
利用本试验设计的金黄色葡萄球菌nuc基因引物和支气管败血波氏杆菌fimN基因引物分别进行PCR扩增。结果显示,扩增得到的目的片段与预期相符,分别为322、478 bp(图 1)。将目的片段纯化回收并测序,测序结果表明扩增得到的2种病原的目的片段与参考序列的同源性均超过99%。
![]() 图 1 单重PCR扩增注:M为DL2000 Marker; 1为金黄色葡萄球菌; 2为支气管败血波氏杆菌。Figure 1. Single PCR amplification
图 1 单重PCR扩增注:M为DL2000 Marker; 1为金黄色葡萄球菌; 2为支气管败血波氏杆菌。Figure 1. Single PCR amplification2.2 双重PCR反应体系的建立及优化
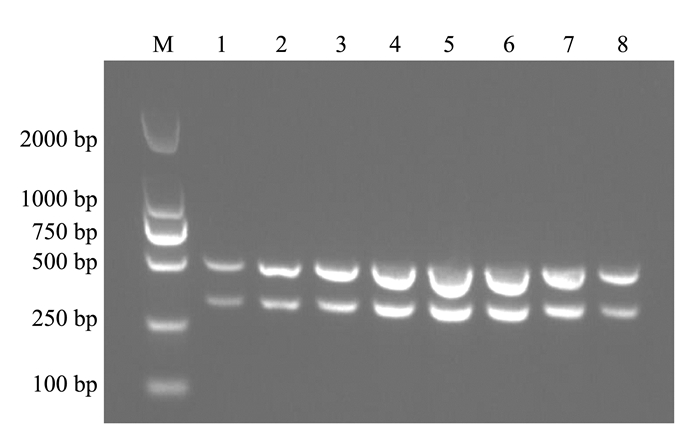
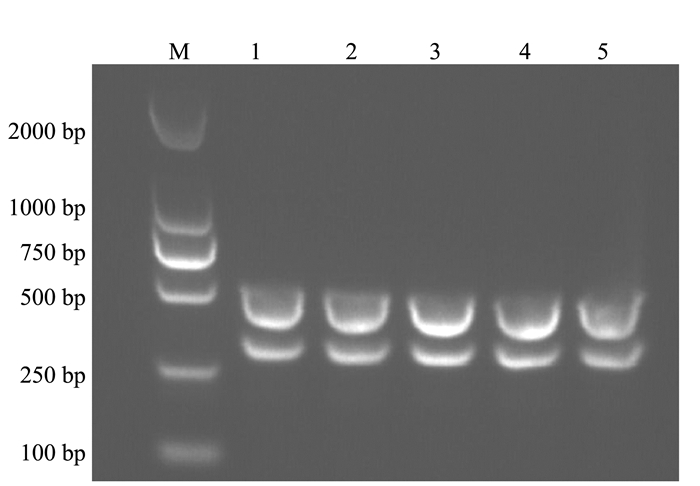
通过固定双重PCR反应体系中模板的量,依次对退火温度和引物浓度进行优化。试验结果表明当退火温度为55.8、57.5和58.8℃(图 2),引物浓度为0.4 μmol·L-1时(图 3),该双重PCR检测方法具有较好的扩增效果。根据退火温度高,特异性强的原则,确定最佳的退火温度为58.8℃。
![]() 图 2 双重PCR退火温度的优化注:M为DL2000 Marker; 1为53℃; 2为53.5℃; 3为54.4℃; 4为55.8℃; 5为57.5℃; 6为58.8℃; 7为59.6℃; 8为60℃。Figure 2. Optimization on annealing temperature for duplex PCR assay
图 2 双重PCR退火温度的优化注:M为DL2000 Marker; 1为53℃; 2为53.5℃; 3为54.4℃; 4为55.8℃; 5为57.5℃; 6为58.8℃; 7为59.6℃; 8为60℃。Figure 2. Optimization on annealing temperature for duplex PCR assay![]() 图 3 双重PCR引物浓度的优化注:M为DL2000 Marker; 1为0.2 μmol·L-1; 2为0.3 μmol·L-1; 3为0.4 μmol·L-1; 4为0.5 μmol·L-1; 5为0.6 μmol·L-1。Figure 3. Optimization on primer concentrations for duplex PCR assay
图 3 双重PCR引物浓度的优化注:M为DL2000 Marker; 1为0.2 μmol·L-1; 2为0.3 μmol·L-1; 3为0.4 μmol·L-1; 4为0.5 μmol·L-1; 5为0.6 μmol·L-1。Figure 3. Optimization on primer concentrations for duplex PCR assay2.3 双重PCR的特异性试验
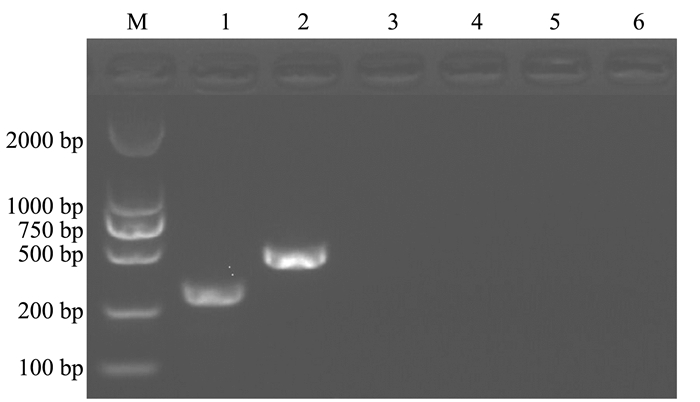
对多杀性巴氏杆菌、大肠杆菌和魏氏梭菌进行检测,结果显示该双重PCR方法与3种常见兔病原菌无交叉反应(图 4),表明该方法具有较好的特异性。
![]() 图 4 双重PCR特异性试验注:M为DL2000 Marker; 1为金黄色葡萄球菌; 2为支气管败血波氏杆菌; 3为多杀性巴氏杆菌; 4为大肠杆菌; 5为魏氏梭菌; 6为灭菌ddH2O。Figure 4. Specificity of duplex PCR assay
图 4 双重PCR特异性试验注:M为DL2000 Marker; 1为金黄色葡萄球菌; 2为支气管败血波氏杆菌; 3为多杀性巴氏杆菌; 4为大肠杆菌; 5为魏氏梭菌; 6为灭菌ddH2O。Figure 4. Specificity of duplex PCR assay2.4 双重PCR的敏感性试验
对反应体系中DNA基因组含量为5 fg~50 ng的金黄色葡萄球菌和支气管败血波氏杆菌进行敏感性试验。当金黄色葡萄球菌和支气管败血波氏杆菌DNA基因组含量均低于5 pg时,扩增的目的条带较不明显(图 5)。结果说明,在50 μL PCR反应体系中,该双重PCR方法能检测到5 pg的金黄色葡萄球菌和支气管败血波氏杆菌DNA基因组。
![]() 图 5 双重PCR敏感性试验注:M为DL2000 Marker; 1为50 ng; 2为5 ng; 3为500 pg; 4为50 pg; 5为5 pg; 6为500 fg; 7为50 fg; 8为5 fg。Figure 5. Sensitivity of duplex PCR assay
图 5 双重PCR敏感性试验注:M为DL2000 Marker; 1为50 ng; 2为5 ng; 3为500 pg; 4为50 pg; 5为5 pg; 6为500 fg; 7为50 fg; 8为5 fg。Figure 5. Sensitivity of duplex PCR assay2.5 临床样品的检测
应用建立的双重PCR方法对63份临床样品进行检测,结果(表 2)表明双重PCR的检测结果与已知结果的符合率为100%。说明该双重PCR方法准确性好。
表 2 临床样品检测结果Table 2. Pathogen detection of duplex PCR assay on clinic samples样品 已知结果/份 双重PCR方法鉴定结果/份 符合率/% 金黄色葡萄球菌 20 20 100 支气管败血波氏杆菌 24 24 100 混合感染 14 14 100 阴性 5 5 100 总计 63 63 100 3. 讨论
在众多引起兔呼吸道疾病的病原中,金黄色葡萄球菌和支气管败血波氏杆菌感染导致的临床症状和病变特征最相似,给这两种病原的鉴别诊断带来困难。金黄色葡萄球菌和支气管败血波氏杆菌的感染率和发病率均较高,且临床上主要依赖抗生素进行防治,药物的频繁使用导致耐药菌株的出现,使疾病愈来愈复杂[5-6, 9]。
nuc基因是金黄色葡萄球菌的特异基因。Brakstad OG等利用该基因上的保守序列建立了检测金黄色葡萄球菌的单重PCR检测方法[10]。向正刚等对四川部分区域分离的41株兔源金黄色葡萄球菌的研究结果表明,所有菌株均携带nuc基因[11]。fimN基因是支气管败血波氏杆菌的菌毛基因,保守、特异,编码的蛋白与细菌毒力相关。刘燕等[12]根据该基因设计了1对特异性引物,建立了检测兔支气管败血波氏杆菌的PCR快速检测方法。此外,还通过原核表达的fimN蛋白建立了间接ELISA方法[7, 13]。综上所述,以金黄色葡萄球菌nuc基因和支气管败血波氏杆菌fimN基因为目的基因能够建立检测这两种病原菌的特异性检测方法。
双重PCR反应体系不是单重PCR反应体系的单纯叠加,反应条件的优化是双重PCR建立的关键。本试验对退火温度和引物浓度进行优化后,确定最佳的退火温度为58.8℃,引物按1:1混合,最佳浓度为0.4 μmol·L-1。该双重PCR方法敏感性高,最低检出极限与刘燕等[12]建立的以fimN基因为目的基因的兔支气管败血波氏杆菌PCR检测方法和凤晓博等[14]建立的以nuc基因为目的基因的金黄色葡萄球菌PCR检测方法相同,能达到pg级;同时,该双重PCR方法特异性好,对其他3种常见兔病原菌无交叉反应。同时,通过对临床样品进行检测,该双重PCR方法与已知结果的符合率为100%。综上所述,本试验建立的双重PCR方法具有良好的特异性、敏感性和准确性,为兔金黄色葡萄球菌和支气管败血波氏杆菌的鉴别诊断提供了有效的技术手段。
-
![]()
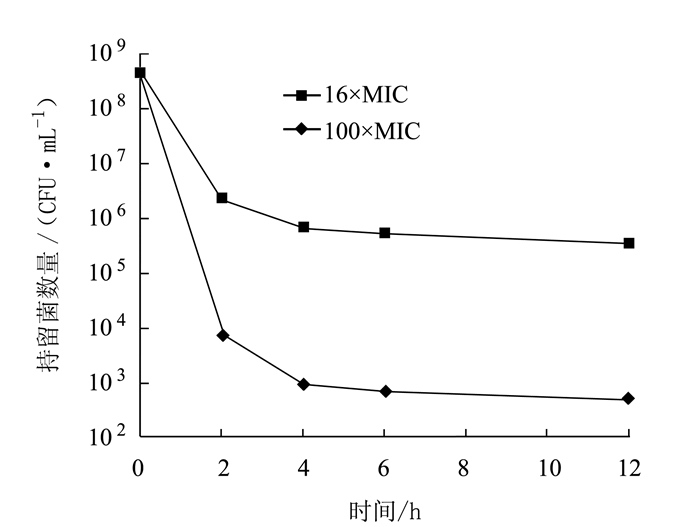
图 1 不同Gen浓度下鸡源奇异变形杆菌PM2658持留菌形成情况
Figure 1. Formation of P. mirabilis PM2658 persisters under various concentrations of Gen
![]()
图 2 鸡源奇异变形杆菌PM2658持留菌遗传学特性检测结果
注:A为未经抗生素处理PM2658和经16×MIC和100×MIC Gen处理4 h后持留菌在不含抗生素培养基上的生长情况;B为经过16×MIC和100×MIC Gen处理4 h后持留菌在含有相应浓度Gen培养基上的生长情况。
Figure 2. Growth of P. mirabilis PM2658 persisters on culture media containing different amount of Gen
![]()
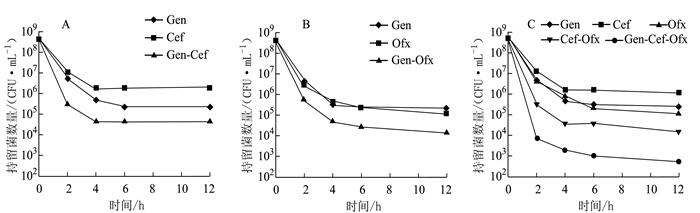
图 3 鸡源奇异变形杆菌持留菌多重抗生素耐受性
注:A为持留菌对Gen-Cef耐受性;B为持留菌对Gen-Ofx耐受性;C为持留菌对Gen-Cef-Ofx耐受性。
Figure 3. Multi-drug tolerance of P. mirabilis PM2658 persisters
-
[1] BIGGER J W. Treatment of staphylococcal infections with penicillin by intermittent sterilization[J]. Lancet, 1944, 244(6320):497-500. DOI: 10.1016/S0140-6736(00)74210-3
[2] COHEN N R, LOBRITZ M A, COLLINS J J. Microbial persistence and the road to drug resistance[J]. Cell Host Microbe, 2013, 13(6):632-642. DOI: 10.1016/j.chom.2013.05.009
[3] ADAMS K N, TAKAKI K, CONNOLLY L E, et al. Drug tolerance in replicating mycobacteria mediated by a macrophage-induced efflux mechanism[J]. Cell, 2011, 145(1):39-53. DOI: 10.1016/j.cell.2011.02.022
[4] CHONG Y P, PARK S J, KIM H S, et al. Persistent Staphylococcus aureus bacteria:a prospective analysis of risk factors, outcomes, and microbiologic and genotypic characteristics of isolates[J]. Medicine (Baltimore), 2013, 92(2):98-108. DOI: 10.1097/MD.0b013e318289ff1e
[5] HOFSTEENGE N, NIMWEGEN E V, SILANDER O K. Quantitative analysis of persister fractions suggests different mechanisms of formation among environmental isolates of E.coli[J]. BMC Microbiology, 2013, 13:25. DOI: 10.1186/1471-2180-13-25
[6] HOLLING N, DEDI C, JONES C E, et al. Evaluation of environmental scanning electron microscopy for analysis of Proteus mirabilis crystalline biofilms in situ on urinary catheters[J]. FEMS Microbiology Letters, 2014, 355(4):20-27. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=JJ0233660810
[7] BORGMAN C J. Proteus mirabilis and its role in dacryocystitis[J]. Optometry and Vision Science, 2014, 91(9):230-235. DOI: 10.1097/OPX.0000000000000347
[8] 周芳, 冉丹丹, 刘飞, 等.鸡源奇异变形杆菌的分离鉴定及系统进化分析[J].动物医学进展, 2015, 36(7):29-32. DOI: 10.3969/j.issn.1007-5038.2015.07.006 [9] 李欣南, 韩镌竹, 宁宜宝.鸡源奇异变形杆菌的分离鉴定及耐药性分析[J].黑龙江畜牧兽医, 2015, (6):165-167, 289. http://www.cnki.com.cn/Article/CJFDTOTAL-HLJX201511055.htm [10] CLINICAL AND LABORATORY STANDARDS INSTITUTE. Reference method for broth dilution antifungal susceptibility testing of yeasts-second edition:approved standard M27-A[M]. Wayne, PA:CLSI, 2002.
[11] BUTT A, HIGMAN V A, WILLIAMS C, et al. The HicA toxin from Burkholderia pseudomallei has a role in persister cell formation[J]. The Biochemical Journal, 2014, 459(2):333-344. DOI: 10.1042/BJ20140073
[12] 胡丽庆, 史煜波, 孙定河, 等.奇异变形杆菌耐药性的4年监测及碳青霉烯类耐药株的耐药机制研究[J].中国微生态学杂志, 2012, 24(7):611-614. http://d.old.wanfangdata.com.cn/Periodical/zgwstxzz201207010 [13] PEARSON M M, SEBAIHIA M, CHURCHER C, et al. Complete genome sequence of uropathogenic Proteus mirabilis, a master of both adherence and motility[J]. Journal of Bacteriology, 2008, 190(11):4027-4037. DOI: 10.1128/JB.01981-07
[14] GRANT S S, HUNG D T. Persistence bacterial infections, antibiotic tolerance, and the oxidative stress response[J]. Virulence, 2013, 4(4):273-283. DOI: 10.4161/viru.23987
[15] GUSAROV I, SHATALIN K, STARODUBTSEVA M, et al. Endogenous nitric oxide protects bacteria against a wide spectrum of antibiotics[J]. Science, 2009, 325(5964):1380-1384. DOI: 10.1126-science.1175439/
[16] LIU Y, IMLAY J A. Cell death from antibiotics without the involvement of reactive oxygen species[J]. Science, 2013, 339(6124):1210-1213. DOI: 10.1126/science.1232751
[17] FENG J, WEITNER M, SHI WL, et al. Eradication of biofilm-like microcolony structures of Borrelia burgdorferi by daunomycin and daptomycin but not mitomycin C in combination with doxycycline and cefuroxime[J]. Front Microbiol, 2016, 7:62. http://pubmedcentralcanada.ca/pmcc/articles/PMC4748043/
[18] PU Y Y, ZHAO Z L, LI Y X, et al. Enhanced efflux activity facilitates drug tolerance in dormant bacterial cells[J]. Mol Cell, 2016, 62(2):284-294. DOI: 10.1016/j.molcel.2016.03.035
[19] BALABAN, N Q, MERRIN J, CHAIT R, et al. Bacterial persistence as a phenotypic switch[J]. Science, 2004, 305(5690):1622-1625. DOI: 10.1126/science.1099390
[20] KORCH S B, HILL T M. Ectopic overexpression of wild-type and mutant hipA genes in Escherichia coli:effects on macromolecular synthesis and persister formation[J]. Journal of Bacteriology, 2006, 188(11):3826-3836. DOI: 10.1128/JB.01740-05
[21] FUNG D K, CHAN E W, CHIN M L. Delineation of a bacterial starvation stress response network which can mediate antibiotic tolerance development[J]. Antimicrobial Agents and Chemotherapy, 2010, 54(3):1082-1093. DOI: 10.1128/AAC.01218-09
[22] NGUYEN D, JOSHI-DATAR A, LEPINE F, et al. Active starvation response mediate antibiotic tolerance in biofilms and nutrient-limited bacteria[J]. Science, 2011, 334(6058):982-986. DOI: 10.1126/science.1211037
-
期刊类型引用(4)
1. 杨帅,段宏伟,吕建树,曾建林,闫振兴,胡俊杰,张勇,赵兴绪. 甘肃省庆阳地区湖羊皮下脓肿病的流行病学调查及病原体分析. 核农学报. 2023(12): 2510-2517 .  百度学术
百度学术
2. 杨鹏,吴燕,岳筠,陈静,李梅,王慧,张双翔,文明,程振涛. 绵羊肺炎支原体P113蛋白C末端基因真核表达载体的构建及其小鼠免疫应答. 中国兽医学报. 2022(03): 496-501+521 . 百度学术
3. 杨鹏,杨源,岳筠,陈静,王慧,朱二鹏,张双翔,文明,程振涛. 绵羊肺炎支原体感染对贵州不同品种山羊肺脏和血液Toll样受体基因转录水平的影响. 动物医学进展. 2022(12): 1-9 . 百度学术
4. 尹峥,刘刚,王晶晶,李晨露,徐海玲,张琪,许信刚. 基于PLD蛋白的伪结核棒状杆菌血清抗体间接ELISA检测方法的建立与应用. 中国兽医科学. 2021(01): 9-16 . 百度学术
其他类型引用(4)
 下载:
下载:

计量
- 文章访问数: 1214
- HTML全文浏览量: 190
- PDF下载量: 10
- 被引次数: 8
