

Construction and Genetic Background Recovery of Blast-resistant Near-isogenic Lines from Japonica Variety, Yunyin
-
摘要: 通过对稻瘟病抗性近等基因系的构建和评价,为抗稻瘟病基因的抗性分析和遗传学研究提供良好载体。分别以广谱抗稻瘟病品种云引和普感稻瘟病品种丽江新团黑谷(LTH)为抗病基因的供体亲本和轮回亲本,结合分子标记辅助选择和人工接种抗性鉴定,经连续4代回交和8代自交,获得7个以LTH为遗传背景的稻瘟病抗性近等基因系。选取均匀分布于水稻12条染色体上的184对SSR标记,对所获得的近等基因系进行遗传背景分析、评价。结果表明:在亲本云引和LTH之间具多态性的SSR引物有72对,标记多态性频率为39.1%。以在亲本间具有多态性的标记对7个近等基因系与LTH间的遗传多态性进行分析,发现其中W15、W17的遗传背景恢复率为97.61%,W18的遗传背景恢复率为97.22%,其他4个株系的遗传背景恢复率均大于98%。成功构建并获得了以LTH为背景的含有云引稻瘟病抗性基因的近等基因系,且近等位基因系的遗传背景与表型值均恢复良好。Abstract: Construction of near-isogenic lines(NILs) is an effective mean for analyzing blast-resistance genes as well as genetic research in general. This study applied the broad-spectrum blast-resistant japonica variety, Yunyin, as the donor parent and the blast-susceptible variety, LTH, as the recurrent parent. Combining the molecular markers assisted selection and artificial inoculation, 7 blast-resistant NILs were bred with LTH for the genetic background analysis after 4 generations of backcrossing and 8 generations of self-crossing.184 pairs of SSR markers evenly distributed on 12 chromosomes of the rice were selected to evaluate the genetic background of the NILs. The results showed that 72 pairs of SSR primers to be polymorphic with a frequency of 39.1%. The polymorphisms between 7 NILs and LTH were analyzed by markers among the parents. The background recovery rates of W15 and W17 were 97.61%, that of W18, 97.22%, and those of other 4 lines, greater than 98%. It appeared that the construction of NILs carrying Yunyin blast-resistance gene with LTH as background were successful, and that all phenotypic indices of the NILs recovered well in this study.
-
Keywords:
- rice /
- rice blast-resistance /
- near-isogenic lines /
- genetic background
-
水稻是全世界最重要的粮食作物之一,全球超过50%的人口以大米为主食[1]。稻瘟病是水稻最主要的病害之一,每年造成水稻产量损失10%~20%,局部地区的损失超过50%甚至绝收[2]。因此,不断从种质资源中挖掘新抗病基因并将其应用于抗病育种及农业生产,一直是水稻育种的重要研究领域。
从抗病基因的发掘、鉴定到生产应用是一个长期和复杂的过程。为了更好地了解抗病基因的抗谱,及其在不同遗传背景下的抗性水平,通常需要综合运用遗传学、基因组学及其他分子生物学的技术对基因的功能及抗病机制进行深入分析,而构建近等基因系是其中常用的遗传学方法之一。近等基因系(Near Isogenic Lines, NILs)的构建是指通过饱和回交,获得理论上除了目标性状有差异之外,其他遗传背景基本都相同的两个遗传材料(品系)[3]。该方法常用于水稻育种中品系的定向改良、功能基因的定位、基因效应分析及稻瘟病菌生理小种遗传群体结构等研究领域[3],在水稻的抗病性遗传改良、抗病基因与病原菌互作的遗传学,以及在分子生物学研究中已得到广泛应用,并取得了良好的效果。章奇等[4]以改良型粳稻品种沈农1033为轮回亲本,通过连续回交,结合各世代的抗性鉴定和农艺性状筛选,转育成3个具有较好应用价值的粳型抗白叶枯病近等基因系。褚晋等[5]分别用LTH、CO39和IRBL系列的稻瘟病抗性近等基因系对云南的稻瘟病菌生理小种结构进行分析,认为Pi-2和Pi-9、Pi-2和Pi-12适合在抗病育种中聚合应用,该研究结果为水稻的抗稻瘟病育种亲本的选择提供了参考。朱小源等[6]利用30个抗稻瘟病的近等基因系对146个来自广东的稻瘟病菌株进行致病性测定,筛选出9个对我国籼稻区稻瘟病菌致病性有较强鉴别力的近等基因系,可用于华南稻区稻瘟病菌生理小种的划分。赵霖等[7]以原始受体亲本秀水11为轮回亲本,外源豌豆铁蛋白转基因纯系Fer34为供体亲本,通过连续回交育成了以秀水11为遗传背景的近等基因系,这为开展富铁水稻新品种的培育提供了依据。水稻品种丽江新团黑谷(LTH)是云南粳型地方品种,该品种对来自全国各地的2 460个、来自日本的200个、来自菲律宾的80个及来自韩国的100个代表性稻瘟病菌株表现高感[8],是用于构建稻瘟病抗性近等基因系的良好载体。
基于前人的研究结果和方法,本研究以广谱稻瘟病抗性品种云引为供体亲本,以LTH为轮回亲本,通过杂交、连续回交、室内分子标记辅助选择和田间稻瘟病抗性鉴定相结合的方法,进行稻瘟病抗性近等基因系的构建和评价,以期为稻瘟病抗性新基因的挖掘和分子作用机制解析提供科学依据。
1. 材料与方法
1.1 供试材料
广谱稻瘟病抗性品种云引由云南省农业科学院粮食作物研究所提供,普感品种LTH及供田间注射接菌用的稻瘟病菌生理小种四川-43均由中国农业科学院作物科学研究所雷财林博士提供。
1.2 试验方法
1.2.1 稻瘟病抗性近等基因系的构建
2007年8月在福建省福州市,以广谱抗稻瘟病粳稻品种云引为抗病基因供体亲本,以普感稻瘟病品种LTH为受体亲本进行杂交,获得杂交种F1代种子,同年于海南三亚福建省南繁育种基地种植F1代,次年3月以其为母本与轮回亲本LTH回交,获得BC1F1代种子, 之后分别于福建省沙县福建省农业科学院水稻研究所育种基地和海南三亚福建省南繁育种基地进行连续回交和自交,获得7个BC4F8代稳定株系。
1.2.2 分子标记辅助选择
同时将近等基因材料和轮回亲本按照固定的规格种植于田间,待移栽苗返青后采集单株叶片,用于室内分子标记辅助选择抗病单株。
使用的连锁的SSR分子标记为RM208,引物序列为:
正向:5'-tctgcaagccttgtctgatg-3'
反向:5'-taagtcgatcattgtgtggacc-3'
PCR扩增的反应条件:95℃预变性5 min、95℃变性30 s、56℃退火30 s、72℃延伸30 s,30个循环,最后72℃延伸1 min。PCR反应体系为10.0 μL,其中:PCR Mix 5 μL,正、反向引物各0.5 μL(10 μmol),DNA模板1 μL (50 ng·μL-1)及ddH2O 3 μL。
1.2.3 抗病性鉴定
将近等基因材料和轮回亲本材料(LTH)按照固定的规格种植于田间,每3个近等基因系小区之间种植1个LTH对照小区,待材料长至分蘖盛期一周前(一般为移栽后25 d左右)适量偏施氮肥,用培养好的新鲜稻瘟病菌孢子悬浮液调节浓度至1×105个·mL-1,用于人工注射接菌,每个单株分别接种3个分蘖并挂牌标记,病情指数最高的一个分蘖代表该株的抗性水平。
1.2.4 农艺性状调查分析
将稳定的近等基因系(BC4F8代)和轮回亲本(LTH)分别种植于福建省沙县福建省农业科学院水稻研究所育种基地,每个小区种植49株(7×7),种植规格为22.3 cm×22.3 cm,3次重复,近等基因系编号后随机种植,每隔3个号种植一区对照(LTH),记载生育期及田间株叶形态表现情况,待完全成熟后测量株高并进行相关农艺性状的考查。
1.2.5 遗传背景恢复率分析
根据已公布的水稻SSR遗传连锁图谱[9],分别在水稻12条染色体上均匀选取136对SSR引物及《国家水稻品种鉴定技术规程》(NY/T1433-2014)[10]中用于水稻品种鉴定的48对SSR引物,先进行供体亲本和轮回亲本间的遗传背景多态性分析,再利用筛选到的在双亲间具有多态性的引物,以轮回亲本LTH为对照,对所构建的稳定的近等基因系材料进行遗传多样性分析。选取的引物由上海生工生物工程有限公司合成。其PCR反应体系为10.0 μL,其中5 μL的PCR Mix,正、反向引物各0.5 μL(10 μmol),DNA模板1 μL (50 ng·μL-1)及3 μL的ddH2O。退火温度参照网站推荐的SSR引物退火温度[11]。SSR扩增的反应程序参照陈锦文等[12]的方法,应用ABI PCR System 9700进行扩增。PCR扩增产物通过8.0%聚丙烯酰胺凝胶电泳进行分离,经Gelred染色后置于凝胶成像系统拍照获取电泳图谱。再根据遗传背景恢复率公式[13]分析后代的遗传背景恢复率。
遗传背景恢复率(G)的计算公式为:
Gg=0.5+0.5(Xg/L) 其中:Gg指g代的遗传背景恢复率;Xg指在回交g代表现为轮回亲本带型的分子标记数量;L指所分析的分子标记数量。
2. 结果与分析
2.1 近等基因系的抗病性鉴定与检测
通过逐代利用稻瘟病菌生理小种四川43对回交后代进行人工注射接菌鉴定,同时以供体亲本云引(高抗)和轮回亲本LTH(高感)分别作为抗病和感病对照。试验结果表明,供体亲本云引表现为对四川43高抗,轮回亲本LTH对四川43表现高感,对自交稳定的7个近等基因系进行人工注射接菌,鉴定结果也均表现为抗病。其中W13、W14、W15、W17、W18、W19、W22为田间表现良好抗性的近等基因系,S1为田间表现高感的株系(表 1)。
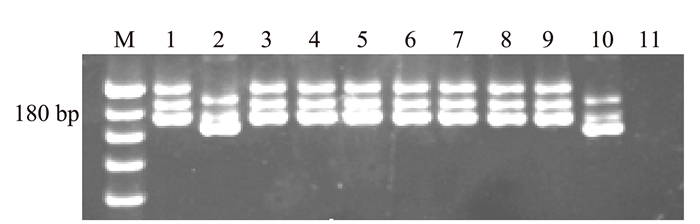
表 1 近等基因系和亲本的人工注射接菌及分子标记检测Table 1. Artificial inoculation and molecular marker detection of NILs and parents品种(系) 注射接菌结果(抗性) 分子检测结果 云引 高抗(HR) + LTH 高感(HS) - W13 抗病(R) + W14 高抗(R) + W15 抗病(R) + W17 抗病(R) + W18 中抗(MR) + W19 中抗(MR) + W22 抗病(R) + S1 高感(HS) - 注:①人工注射接菌时对接种叶片造成了伤口,病情略微偏重,本研究将调查结果中抗及以上的均记为抗病表型;②+表示阳性,-表示阴性。 在进行人工注射接菌鉴定之后,对接种的单株分别取叶片对目标基因进行分子标记检测的结果(表 1)表明,本研究筛选出的抗病近等基因系材料均能扩增出与供体亲本云引相同的目的条带(图 1)。由此可见,本试验已经成功地将云引所携带的广谱稻瘟病抗性基因导入到LTH的遗传背景之中,使其获得了与云引一样的抗稻瘟病菌株(四川43)侵染的能力。
![]() 图 1 近等基因系中目标基因分子标记检测注:M为20bp DNA marker;1为供体亲本云引;2为轮回亲本LTH;3~9为田间接种抗病近等基因系,对应编号为:W13、W14、W15、W17、W18、W19、W22;10为感病近等基因系S1;11为阴性对照(水)。Figure 1. Detection of molecular markers of target genes in NILs
图 1 近等基因系中目标基因分子标记检测注:M为20bp DNA marker;1为供体亲本云引;2为轮回亲本LTH;3~9为田间接种抗病近等基因系,对应编号为:W13、W14、W15、W17、W18、W19、W22;10为感病近等基因系S1;11为阴性对照(水)。Figure 1. Detection of molecular markers of target genes in NILs2.2 近等基因系的农艺性状
分别在水稻齐穗后30 d,对供体亲本、受体亲本及近等基因系材料各取10个单株进行株高、分蘖数、穗粒数、穗长、结实率、千粒重等主要农艺性状和产量性状的数据进行测量和调查,统计结果见表 2。
表 2 供体亲本、受体亲本及近等基因系材料的主要农艺性状Table 2. Agronomic traits of donor parents, receptor parents and NILs品种(系) 株高/cm 分蘖数/个 穗长/cm 株总粒数/粒 株实粒数/粒 穗总粒数/粒 穗实粒数/粒 结实率/% 千粒重/g 云引 128.6 12.7 22.4 1967.3 1678.0 154.6 131.9 85.3 29.3 LTH 134.7 18.3 21.1 2584.6 2149.7 141.0 117.2 83.1 26.6 W13 133.2 17.6 22.2 2306.8 1923.4 130.5 108.8 83.3 27.8 W14 134.5 19.3 23.0 2798.6 2302.2 144.7 119.1 82.2 27.2 W15 131.2 18.6 20.5 2524.6 2083.8 135.2 111.6 82.5 26.7 W17 135.4 17.8 20.8 2396.3 2009.0 133.9 112.3 83.8 27.1 W18 131.9 18.3 21.4 2448.6 2035.7 133.5 111.0 83.1 26.9 W19 135.8 17.9 22.5 2502.9 2062.5 139.5 115.0 82.4 27.0 W22 130.9 18.6 21.1 2675.0 2209.9 143.2 118.3 82.6 27.1 试验结果表明,本研究利用分子标记辅助选择和人工注射接种的方法获得的携带云引稻瘟病抗性基因的7个近等基因系材料与受体亲本LTH在上述农艺性状上均没有明显差异(图 2),该近等基因系是应用蛋白质组学、基因组学等方法对抗病的目标基因进行系统研究的优异材料。
![]() 图 2 供体亲本云引、受体亲本LTH及其近等基因系W22的株型表现Figure 2. Plant types of donor parent Yunyin, recipient parent LTH and their NIL W22
图 2 供体亲本云引、受体亲本LTH及其近等基因系W22的株型表现Figure 2. Plant types of donor parent Yunyin, recipient parent LTH and their NIL W222.3 近等基因系的遗传背景恢复率
根据水稻数据库中公布的SSR标记[9],用均匀分布在水稻12条染色体上、平均间隔10 cm的136对SSR引物及《国家水稻品种鉴定技术规程》(NY/T 1433-2014)[10]中用于品种指纹图谱分析的48对SSR引物,分别对供体亲本和受体亲本进行多态性标记的筛选。在双亲间共获得72对多态性标记,多态性频率为39.1%。本研究中多态性引物选择的标准为:①扩增产物在8%聚丙烯酰胺凝胶电泳上的带型清晰可辨,特异性好;②在供体亲本和轮回亲本之间有不同的等位变异,位点之间仅有1~2个bp差异的引物予以淘汰。选择在亲本间具有明显多态性且带型清晰可辨的标记,对育成的7个BC4F8代株系进行遗传背景恢复率分析(图 3)。统计结果表明,这7个株系的基因型均恢复到轮回亲本LTH基因型的97%以上。其中株系W15、W17的遗传背景恢复率为97.61%,W18的遗传背景恢复率为97.22%,其他4个株系的遗传背景恢复率均大于98%(表 3)。且所有多态性的标记均包含了本研究中用于抗性基因检测的标记RM208。本研究成功构建并获得了以LTH为背景的抗稻瘟病近等基因系,且近等基因系的遗传背景与表型值均恢复良好,农艺性状与生育期均表现一致,可供后续蛋白质组学、基因功能与抗谱分析等研究。
![]() 图 3 用SSR标记对近等基因系进行遗传背景分析注:M为20 bp marker,A1~A12、B1、B2均为水稻品种鉴定技术规程(NY/T 1433-2014)中的引物。样品顺序为LTH、云引、分子标记为阳性株、分子标记为阴性株。Figure 3. Genetic background analysis of NILs by SSR markers表 3 近等基因系与轮回亲本LTH之间的多态性标记及其遗传背景恢复率Table 3. Genetic background recovery rates and polymorphic markers between NILs and LTH
图 3 用SSR标记对近等基因系进行遗传背景分析注:M为20 bp marker,A1~A12、B1、B2均为水稻品种鉴定技术规程(NY/T 1433-2014)中的引物。样品顺序为LTH、云引、分子标记为阳性株、分子标记为阴性株。Figure 3. Genetic background analysis of NILs by SSR markers表 3 近等基因系与轮回亲本LTH之间的多态性标记及其遗传背景恢复率Table 3. Genetic background recovery rates and polymorphic markers between NILs and LTH品系 多态性标记数 恢复率/% W13 2 98.61 W14 2 98.61 W15 3 97.61 W17 3 97.61 W18 4 97.22 W19 1 99.31 W22 1 99.31 3. 讨论与结论
近等基因系的构建是研究未克隆基因遗传学特性的有效手段之一,也是在水稻育种中常用的方法。除了肉眼可见的农艺性状的一致性之外,近等基因系构建的优劣主要取决于其遗传背景的恢复率,遗传背景恢复率高的近等基因系用于后续研究时可以在较大程度上排除其他非目标基因的干扰,有针对性地对目的基因的功能和作用机制进行系统研究。要获得高恢复率的近等基因系,除了通过系统的表型调查和抗性鉴定等来判断之外,还需通过采用分子标记手段从全基因组水平进行筛选和判断。分子标记辅助选择是利用与目标基因紧密连锁或基因本身的分子标记来进行目标基因的选择,这种方法可在水稻的全生育期进行,不受环境或植株状态的影响,被广泛应用于对遗传材料的遗传背景恢复率的检测。沈新莲等[14]的综述结果表明,对目标基因和理想状态的遗传背景进行选择时,分子标记辅助选择是有效的,在回交二代后,分子标记的跨度为10~20 cM较适宜,这为利用适合距离的分子标记进行遗传背景的选择提供了理论依据。通过一代回交,理论上能够替换轮回亲本50%的基因片段,回交一代的种子占轮回亲本的75%,回交二代种子占轮回亲本的87.5%,依此类推。Xu等[15]的研究结果表明,对多数物种来说,有适合数量的标记和足够的后代用于背景恢复率选择,两代以内即可恢复到轮回亲本基因型的90%左右。本研究选择均匀分布于水稻12条染色体上的184对SSR引物,平均遗传距离约为10 cM,在各个回交和自交世代对目标基因进行跟踪选择,并通过田间接种鉴定进行表型选择,结合单株田间农艺性状的筛选进行4代回交和8代自交。利用在供体亲本和轮回亲本间具有多态性的40对SSR引物进行分析,最终获得7个遗传背景恢复率在97%以上的近等基因系。本研究结果表明,用分子标记辅助选择与表型选择相叠加的方法可提高品种选育的效率,选择遗传背景差异较小的亲本作为供体,对于加快回交导入、提高育种效率有较大的帮助。
此外,分子标记辅助选择也是水稻品种定向改良的有效手段。利用分子标记辅助选择已选育出单一抗性或多基因聚合的品种[16]。郑康乐等的研究结果表明,为提高选择效率,用于分子标记辅助选择的引物与基因之间的遗传距离要小于5 cM,且距离越近检测的准确率越高[17]。本研究运用的与目标基因连锁的分子标记RM208与目标基因的遗传距离为2.6 cM,为本研究中稳定可靠的近等基因系的获得提供了保障。获得了以普感稻瘟病品种LTH为遗传背景,携带广谱稻瘟病抗性品种云引抗病基因的近等基因系,为该抗病基因的后续研究提供了可靠的遗传材料。但另一方面,本研究中的供体亲本和受体亲本均为云南地方粳稻品种,本身遗传多态性不高,致使用于筛选的多态性引物最终只有40对被选作用于遗传背景的恢复率分析,这在用于近等基因系的遗传背景分析时数目偏少,今后希望通过增加标记数量,以保障获得的近等基因系的遗传背景的一致性。
-
![]()
图 1 近等基因系中目标基因分子标记检测
注:M为20bp DNA marker;1为供体亲本云引;2为轮回亲本LTH;3~9为田间接种抗病近等基因系,对应编号为:W13、W14、W15、W17、W18、W19、W22;10为感病近等基因系S1;11为阴性对照(水)。
Figure 1. Detection of molecular markers of target genes in NILs
![]()
图 2 供体亲本云引、受体亲本LTH及其近等基因系W22的株型表现
Figure 2. Plant types of donor parent Yunyin, recipient parent LTH and their NIL W22
![]()
图 3 用SSR标记对近等基因系进行遗传背景分析
注:M为20 bp marker,A1~A12、B1、B2均为水稻品种鉴定技术规程(NY/T 1433-2014)中的引物。样品顺序为LTH、云引、分子标记为阳性株、分子标记为阴性株。
Figure 3. Genetic background analysis of NILs by SSR markers
表 1 近等基因系和亲本的人工注射接菌及分子标记检测
Table 1 Artificial inoculation and molecular marker detection of NILs and parents
品种(系) 注射接菌结果(抗性) 分子检测结果 云引 高抗(HR) + LTH 高感(HS) - W13 抗病(R) + W14 高抗(R) + W15 抗病(R) + W17 抗病(R) + W18 中抗(MR) + W19 中抗(MR) + W22 抗病(R) + S1 高感(HS) - 注:①人工注射接菌时对接种叶片造成了伤口,病情略微偏重,本研究将调查结果中抗及以上的均记为抗病表型;②+表示阳性,-表示阴性。  下载: 导出CSV
下载: 导出CSV
表 2 供体亲本、受体亲本及近等基因系材料的主要农艺性状
Table 2 Agronomic traits of donor parents, receptor parents and NILs
品种(系) 株高/cm 分蘖数/个 穗长/cm 株总粒数/粒 株实粒数/粒 穗总粒数/粒 穗实粒数/粒 结实率/% 千粒重/g 云引 128.6 12.7 22.4 1967.3 1678.0 154.6 131.9 85.3 29.3 LTH 134.7 18.3 21.1 2584.6 2149.7 141.0 117.2 83.1 26.6 W13 133.2 17.6 22.2 2306.8 1923.4 130.5 108.8 83.3 27.8 W14 134.5 19.3 23.0 2798.6 2302.2 144.7 119.1 82.2 27.2 W15 131.2 18.6 20.5 2524.6 2083.8 135.2 111.6 82.5 26.7 W17 135.4 17.8 20.8 2396.3 2009.0 133.9 112.3 83.8 27.1 W18 131.9 18.3 21.4 2448.6 2035.7 133.5 111.0 83.1 26.9 W19 135.8 17.9 22.5 2502.9 2062.5 139.5 115.0 82.4 27.0 W22 130.9 18.6 21.1 2675.0 2209.9 143.2 118.3 82.6 27.1
下载: 导出CSV
表 3 近等基因系与轮回亲本LTH之间的多态性标记及其遗传背景恢复率
Table 3 Genetic background recovery rates and polymorphic markers between NILs and LTH
品系 多态性标记数 恢复率/% W13 2 98.61 W14 2 98.61 W15 3 97.61 W17 3 97.61 W18 4 97.22 W19 1 99.31 W22 1 99.31
下载: 导出CSV
-
[1] 薛恒.米业产业化经营:米业产品的营销创新[J].中国稻米, 2003, 9(5):28-29. DOI: 10.3969/j.issn.1006-8082.2003.05.016 [2] 刘萌.培育水稻空育131BL多系品种供体亲本抗稻瘟病性鉴定及目的基因分子标记分析[D].哈尔滨: 黑龙江大学, 2012. [3] 马玉银, 左示敏, 张再金, 等.水稻近等基因系构建及其应用[J].安徽农业科学, 2008, 36(17):7167-7168. DOI: 10.3969/j.issn.0517-6611.2008.17.049 [4] 章琦, 杨文才, 施爱农, 等. 3个粳稻抗白叶枯病近等基因系的构建[J].作物学报, 1998, 24(6):799-804. DOI: 10.3321/j.issn:0496-3490.1998.06.026 [5] 褚晋, 何鹏飞, 赵正龙, 等. 3套水稻近等基因系对两个病圃稻瘟病菌的抗性评价[J].江西农业学报, 2014, 26(7):50-54. DOI: 10.3969/j.issn.1001-8581.2014.07.014 [6] 朱小源, 杨祁云, 杨健源, 等.抗稻瘟病单基因系对籼稻稻瘟病菌小种鉴别力分析[J].植物病理学报, 2004, 34(4):361-368. DOI: 10.3321/j.issn:0412-0914.2004.04.013 [7] 赵霏, 任三娟, 郭泽建, 等.利用近等基因系研究豌豆铁蛋白基因对水稻重要生物学特性的影响[J].农业生物技术学报, 2011, 19(1):63-68. DOI: 10.3969/j.issn.1674-7968.2011.01.009 [8] 凌忠专, 蒋琬如, 王久林, 等.水稻品种丽江新团黑谷普感特性的研究和利用[J].中国农业科学, 2001, 34(1):116-116. DOI: 10.3321/j.issn:0578-1752.2001.01.025 [9] TEMNYKH S, DeCLERK G, LUKASHOVA A, et al. Computational and experimental analysis of microsatellites in rice (Oryza sativa L.):frequency, length variation, transposon associations, and genetic marker potential[J]. Genome Research, 2001, 11(8):1441-1452. DOI: 10.1101/gr.184001
[10] 中华人民共和国农业部.水稻品种鉴定技术规程SSR标记法: NY/T 1433-2014[S].北京: 中国标准出版社, 2014: 9-13. [11] Cold Spring Harbor Laboratory, Oregon State University, EMBL-EBI. SSR Markers Resource-Microsatellite Markers[R/OL].(2016-08-26)[2018-06-13].http://www.gramene.org/microsat/ssr.html
[12] 陈锦文, 朱永生, 张建福, 等.四个杂交水稻骨干恢复系的多态性分析[J].福建农业学报, 2014, 29(4):319-323. DOI: 10.3969/j.issn.1008-0384.2014.04.005 [13] 阳海宁, 韦绍丽, 李孝琼, 等.标记辅助培育水稻抗稻褐飞虱和稻白叶枯病基因聚合系[J].分子植物育种, 2010, 8(1):11-19. http://d.old.wanfangdata.com.cn/Periodical/fzzwyz201001004 [14] 沈新莲, 张天真.作物分子标记辅助选择育种研究的进展与展望[J].高技术通讯, 2003, 13(2):105-110. DOI: 10.3321/j.issn:1002-0470.2003.02.023 [15] XU Y, CROUCH J H. Marker-assisted selection in plant breeding from publication to practice[J]. Crop science, 2008, 48(2):391-407. DOI: 10.2135/cropsci2007.04.0191
[16] LUO Y C, SANGHA J S, WANG S H, et al. Marker-assisted breeding of Xa4, Xa21 and Xa27 in the restorer lines of hybrid rice for broad-spectrum and enhanced disease resistance to bacterial blight[J]. Molecular Breeding, 2012, 30(4):1601-1610. DOI: 10.1007/s11032-012-9742-7
[17] 郑康乐, 黄宁.标记辅助选择在水稻改良中的应用前景[J].遗传, 1997, 19(2):40-44. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK199700879646 -
期刊类型引用(5)
1. 张茜,李登,叶志成,陈明高,张盼. 基于“水-能-碳”关联系统的城镇污水处理厂碳排放研究. 安徽化工. 2024(03): 113-116+120 .  百度学术
百度学术
2. 王红瑞,李晓军,张力,王力萍,姜欣. 水-能源-碳排放复杂关系研究进展及展望. 南水北调与水利科技(中英文). 2023(01): 13-21 . 百度学术
3. 魏心雨,王雁杰,孙梦媛,王智芳,周凯. 基于土地利用结构变化的河南省碳排放时空格局及驱动力分析. 农业现代化研究. 2023(05): 881-891 . 百度学术
4. 马肖迪,谭晓波. 基于LMDI模型的湖南省土地利用碳排放影响因素研究. 湖南工业大学学报. 2019(06): 60-66 . 百度学术
5. 赵先超,马肖迪,胡艺觉. 基于PSR模型的湖南省土地利用碳排放安全评估. 福建农业学报. 2018(08): 828-834 . 本站查看
其他类型引用(4)


计量
- 文章访问数: 1609
- HTML全文浏览量: 221
- PDF下载量: 24
- 被引次数: 9
