

Phenotypic Diversity on Inflorescence of Macadamia spp. Germplasms
-
摘要:目的 对澳洲坚果种质资源花序表型性状进行多样性分析,为澳洲坚果种质资源的鉴定评价以及创新利用提供参考。方法 花序表型性状的观测和描述方法参照《澳洲坚果种质资源描述规范和数据标准》和《澳洲坚果种质资源鉴定技术规范》,数据统计分析应用聚类和主成分分析的方法。结果 40 份供试澳洲坚果种质资源中 90% 种质小花为乳白色、50% 种质小花自花序轴基部向顶端依次开放、77.5% 种质无批次开花;花序长度的变异系数最大为 26.30%,小花长度的变异系数最小为 8.60%。聚类分析将 40 份种质资源在欧氏距离为 4.79 时分为 2 个组群,组群内的种质资源以花序长度和小花数量聚类;主成分分析结果将 6 个表型性状简化为 3 个主成分(花量、花色、开花顺序因子),解释的总变异为 71.752%,更为直观地展现了花序表型特点,其结果与聚类分析基本一致。结论 澳洲坚果种质资源的花序表型性状存在丰富的多样性,花序长度、小花数量、小花颜色和小花开放顺序是花序表型性状多样性构成的主导因子。Abstract:Objective Phenotypic diversity of inflorescence from a collection of 40 Macadamia spp. was determined to evaluate and better utilize the resources.Method Using the “Standards for Description and Data Analysis on Macadamia” and the “Techniques and Codes for Evaluating Macadamia Germplasms” as references, this study was conducted. Data were subjected to cluster and principal component analyses.Result The inflorescence characteristics of the plants showed that 90% of the germplasms had creamy white flowerets, 50% of the flowerets opened sequentially from the base to the top of inflorescence axis, and 77.5% of them did not flower in batch. The maximum coefficient of variation on inflorescence length was 26.30%, and the minimum 8.60%. The cluster analysis based on 6 phenotypic characteristics of the inflorescence divided the 40 germplasms into two groups at an euclidean distance of 4.79. The categorization was mostly determined by the inflorescence length and floweret count. The 6 phenotypic characteristics composed of 3 independent principal components (i.e., floweret count, floweret color, and flowering sequence) constituting 71.752% of the total variance, The principal component analysis directly demonstrated the phenotypic characteristics of inflorescence which basically agreed with the results obtained by the cluster analysis.Conclusion The phenotypic diversity of macadamia inflorescence was abundant among the germplasms. The inflorescence length, floweret count, floweret color, and opening order of floweret were the dominant factors constituted to the diversity on the inflorescence.
-
Keywords:
- Macadamia spp. /
- germplasms /
- inflorescence /
- phenotypic characteristics /
- diversity
-
0. 引言
【研究意义】玉米(Zea mays L.)是我国的主要粮食作物之一[1],粮饲兼用,也是重要的工业原料,在国家粮食安全保障体系中占有举足轻重的地位[2]。随着国家对知识产权保护范围的拓宽和对科技人才的不断重视,育种人对品种权的保护意识也在不断增强[3]。再者,玉米种质资源的有效挖掘利用、分子辅助选育、基因工程等生物新技术在玉米育种过程中的应用,使得玉米育种年限不断缩短,近年申请品种审定和品种权保护的玉米新品种数量也在急剧上升[4]。植物新品种审定、登记和保护制度是农业知识产权中最重要的一种制度,在鼓励育种创新、丰富我国种质资源、保障粮食安全生产等方面有重要的推动作用[5−6]。然而,无论是申请品种审定还是品种权保护,在其实质性审查时都要进行特异性(distinctness)、一致性(uniformity)和稳定性(stability)测试(简称DUS 测试)[7−8]。当前,玉米DUS测试是参照《植物品种特异性、一致性和稳定性测试指南 玉米》(GB/T 19557.24—2018)[7](以下简称《指南》)的要求开展。《指南》中要求测试的数量性状,一般受环境条件影响较大,在田间种植试验中通常需要设置一些标准品种,以矫正环境变化对数量性状表达的影响[8]。《指南》属于国家标准,在全国范围内流通使用。但经过标准品种的种植,发现《指南》中提供的标准品种在云南生态区域生长不太适宜,导致标准品种示例的性状表达状态在云南低纬高原生态区域存在不同程度变化,出现标准品种不标准等问题,限制了玉米DUS测试工作有序开展,影响测试结果可靠性和科学性[9−10]。【前人研究进展】李娟等[11]基于DUS测试,对玉米标准品种在贵州地区的表达差异及多样性进行分析,建议可将用于研究的36个标准品种调减至31个。刘迪发等[12]对玉米DUS测试标准品种在海南性状表达差异性进行分析,结果表明不同标准品种、不同性状在海南的表现与《指南》赋值的差异程度不同。徐振江等[13]对玉米DUS测试标准品种在华南亚热带生态区的表现与优化进行研究,研究结果认为存在一些标准品种的性状表达与《指南》赋值的符合系数不高。任丽等[14]用符合系数对标准品种在上海地区的性状表现与水稻测试指南中的代码进行差异性分析,并进一步对部分性状的标准品种进行了优化。徐振江等[15]对水稻新品种DUS测试目测性状标准品种进行筛选优化,筛选出16个标准品种就已涵盖原有的23个标准品种代表的性状,大大减少了标准品种数量。赵连佳等[16]对红花(Carthamus tinctorius L.)测试指南中的32份标准品种42个相关性状进行调查分析,结果表明大部分红花标准品种及性状在新疆的表现与测试指南相符。Pérez等[17]在不同环境下对13种板栗(Castanea mollissima BL.)性状进行测试筛,选出较稳定性显著性强的5个性状以确定DUS试验中使用的不同描述符的性质。数量性状的表达存在不同程度的差异,LIU等[18]以亚洲毛茛(Ranunculus japonicus Thunb.)DUS测试指南的制定为例,应用统计方法研究DUS测试数量性状筛选、分级以及标准品种选择。【本研究切入点】已报道的相关作物研究大多集中在对标准品种在不同区域性状表达的稳定性和多样性进行研究分析[19],对标准品种进行优化或筛选出更合适标准品种的报道很少,特别是对《指南》中玉米标准品种筛选优化以及有关低纬高原生态区域玉米DUS测试标准品种筛选的研究几乎没有报道。因此,筛选适合在低纬高原生态区域种植的玉米DUS测试标准品种迫在眉睫。【拟解决的关键问题】本研究以80个杂交玉米品种和20个玉米自交系为研究材料,参照《指南》[7]中的要求和《植物品种特异性 一致性 稳定性测试总论》[8](以下简称《总论》)中标准品种筛选原则,筛选出一套适合于云南等低纬高原区域种植的玉米DUS测试标准品种,并对筛选的标准品种进行聚类分析,为筛选适合在低纬高原区域种植的玉米标准品种等相关研究奠定理论基础。
1. 材料与方法
1.1 试验材料
本试验种植了100个适合在低纬高原生态区域生长的玉米品种,包括80个杂交玉米品种和20个玉米自交系。考虑到品种的代表性和尽可能选择遗传背景差异大的品种,从30家不同育种公司和单位收集品种,每家收集了2~5个品种。试验材料具体信息如表1所示。
表 1 100个试验材料品种名称及品种类型Table 1. Names and types of 100 maize varieties tested品种编号
Varieties
number品种名称
Varieties
name品种类型
Varieties
types品种编号
Varieties
number品种名称
Varieties
name品种类型
Varieties
types品种编号
Varieties
number品种名称
Varieties
name品种类型
Varieties
types品种编号
Varieties
number品种名称
Varieties
name品种类型
Varieties
types1 AA323 CS 26 恩白玉9号 CH 51 洱之玉918 CH 76 川单99 CH 2 AA128 CS 27 龙白玉2号 CH 52 富滇8号 CH 77 荣玉99 CH 3 AA235 CS 28 龙白玉999 CH 53 澄白2号 CH 78 弥玉369 CH 4 A001 CS 29 隆白玉3号 CH 54 秋庆402 CH 79 万丰玉 3612 CH 5 S01 CS 30 龙白玉3号 CH 55 砚白1号 CH 80 奎单828 CH 6 S02 CS 31 贵白玉2号 CH 56 凉都玉1号 CH 81 锋玉217 CH 7 S03 CS 32 贵白玉1号 CH 57 林新13号 CH 82 万丰玉939 CH 8 DT006 CS 33 弥玉129 CH 58 宣农玉7号 CH 83 云谷99 CH 9 DT12832 CS 34 腾丰玉198 CH 59 靖黄7号 CH 84 瑞喜731 CH 10 DT33631 CS 35 澄白2号 CH 60 云瑞 8311 CH 85 顶立313 CH 11 DT3363 CS 36 兆白99 CH 61 弥玉002 CH 86 滇喜518 CH 12 BC57 CS 37 隆白玉109 CH 62 云瑞909 CH 87 金黄10号 CH 13 TRWX-9 WS 38 古白玉1号 CH 63 云瑞208 CH 88 瑞黑糯1号 WH 14 DT5133 CS 39 靖青贮1号 CH 64 田单60 CH 89 云黄13号 CH 15 DT37465 CS 40 靖白1号 CH 65 洱之玉666 CH 90 亲明玉1号 CH 16 DT1461 CS 41 瑞佳甜4号 SH 66 珍谷 68 CH 91 富好2号 CH 17 DT37466 CS 42 云甜玉12号 SH 67 云瑞202 CH 92 水金玉211 CH 18 DT1581 CS 43 花超99 SH 68 金滇 1588 CH 93 足玉817 CH 19 DT6413212 CS 44 双色年华 SH 69 宣晟5号 CH 94 胜玉209 CH 20 DT43774 CS 45 毫发520 SH 70 禾圣707 CH 95 迈单520 CH 21 宣成白1号 CH 46 云甜玉11号 SH 71 禾圣668 CH 96 红玉618 CH 22 神禾788 CH 47 瑞佳甜3号 SH 72 易金273 CH 97 红玉711 CH 23 澄白3号 CH 48 玉研212 CH 73 云良10号 CH 98 高农玉203 CH 24 宣白玉10号 CH 49 子玉68 CH 74 渝单958 CH 99 雅玉2号 CH 25 澄白1号 CH 50 科元玉518 CH 75 西南玉99 CH 100 瑞甜7号 SH 表中字母“CS、WS、CH、SH、WH”分别表示“普通玉米自交系、糯质玉米自交系、普通玉米杂交种、甜玉米杂交种、糯质玉米杂交种”。
CS: common maize inbred lines; WS: waxy maize inbred line; CH: common maize hybrid; SH: sweet maize hybrid; and WH: waxy maize hybrid.1.2 试验方法
1.2.1 田间形态性状测试
田间试验设计与管理:田间测试于2022年4月至2023年11月在农业农村部植物新品种DUS测试(昆明)分中心嵩明基地进行两个周期的形态性状测试。地块平整,肥力中等偏上。试验地地理坐标为25.18° N,103.25° E,海拔为
1900 m,年均气温15 ℃左右,年降雨量在1200 ~1400 mm,年平均无霜期约235 d。试验采用随机区组设计,2次重复,塘播,行距75 cm,塘距35 cm,定苗时每穴留2株,4行区。四周设保护行,常规管理同大田一致。本试验材料于2022年4月29日播种,5月22日进行间苗、定苗。整个生育期除草2次,防病虫害喷洒药3次,中耕培土、起垄各1次。1.2.2 性状观测与判别
性状调查方法、时期、部位及判别严格按照《指南》[7]中的具体要求进行,共测试了100个试验材料的37个数量性状(表2)。
表 2 37个数量性状编号及名称Table 2. Codes and names of 37 quantitative characteristics性状编号
Characters
number性状名称
Characters name性状编号
Character
number性状名称
Characters name性状编号
Characters
number性状名称
Characters name2 幼苗:第一叶鞘花青甙显色强度 16 雄穗:侧枝弯曲程度 29 果穗:穗柄长度 3 叶片:绿色程度 17 雄穗:最低位侧枝以上主轴长度 30 果穗:长度 4 散粉期 18 *雄穗:最高位侧枝以上主轴长度 31 果穗:直径 5 抽丝期 19 雄穗:一级侧枝数目 32 果穗:穗行数 6 植株:上部叶片与茎秆夹角 20 雄穗:侧枝长度 33 果穗:形状 7 叶片:弯曲程度 21 茎秆:“之”字型程度 35 仅适用于甜玉米:籽粒黄色程度 9 雌穗:花丝花青甙显色强度 22 茎秆:支持根花青甙显色强度 36 仅适用于甜玉米:籽粒长度 10 雄穗:花药花青甙显色强度 23 叶片:长度 37 仅适用于甜玉米:籽粒宽度 11 雄穗:小穗密度 24 叶片:宽度 38 仅适用于甜玉米:籽粒皱缩程度 12 雄穗:颖片基部花青甙显色强度 25 叶:叶鞘花青甙显色强度 39 籽粒:类型 13 雄穗:颖尖花青甙显色强度 26 植株:穗位高度 43 穗轴:颖片花青甙显色强度 14 雄穗:颖片中部花青甙显色强度 27 植株:高度 15 雄穗:侧枝与主轴夹角 28 植株:穗位高与株高比率 1.2.3 玉米标准品种筛选
玉米标准品种筛选遵循《总论》中标准品种选择的原则进行[8],主要要求标准品种样品要易获得,性状表达的波动性不大,性状表达的变异范围尽可能覆盖,标准品种的数量尽可能少,必要时需要征求相关专家的意见。获得100个试验材料的37个形态性状数据后,进行标准品种筛选时对《指南》[7]中有9个表达状态的性状,遵循筛选的标准品种尽可能优先覆盖代码为1、3、5、7、9对应的状态;对有5个表达状态的性状,标准品种尽可能优先覆盖代码为2、4对应的状态;对有4个表达状态的性状,标准品种尽可能优先覆盖代码为2、3对应的状态。在这样的原则下,筛选的标准品种在满足能充分发挥示例作用下,可以最大限度减少所需要的标准品种数量,从而减少测试成本。在标准品种确定时,通常将符合某个性状某种表达状态的少数品种优先作为候选标准品种。再根据需要的目标性状表达状态逐一增加候选品种数量,最终再优化品种数量,删除冗余品种。
1.2.4 玉米标准品种聚类分析
综合两年的测试数据,以筛选的标准品种形态性状数据为基础,目测性状直接使用观测值,对于测量数据需先转化成代码值再进行分析。利用NTSYSpc2.10e统计软件中similarity程序计算相似系数,以Clustering程序中SHAN进行非加权组平均法(unweighted pair group method with arithmetic mean, UPGMA)聚类分析[20],绘制玉米品种的遗传聚类系统树。
2. 结果与分析
2.1 37个性状描述结果
由表3、4中性状描述的最大值、最小值及覆盖范围可知,研究材料在37个性状的绝大多数表达状态均有分布,但在极少数性状的个别表达状态上没有分布。结果表明,研究材料的来源还不够丰富,一定程度上也说明个别性状的表达状态设置存在不合理性,实际育种和生产中并不存在那么多的表达状态。由表3、4可知,37个性状的变异系数在5.70%~110.89%。其中,变异系数在15%~20%的性状有5个,小于15%的性状有10个,其他性状的变异系数均大于20%,说明大多数性状上获得的数据离散程度较高,100个研究材料仍然具有代表性。
表 3 基于研究材料的目测性状描述结果Table 3. Descriptions on appearances of maize under study性状编号
Characters
number最大值
Max最小值
Min覆盖范围
Coverage
area变异系数
Coefficient of variation/
%2 8 1 1~8 27.69 3 3 2 2~3 20.41 4 9 1 1~9 24.98 5 9 1 1~9 25.08 6 6 2 2~6 23.89 7 6 1 1~6 33.73 9 7 1 1~7 56.57 10 7 1 1~7 42.86 11 5 2 2~5 18.88 12 8 1 1~8 71.48 13 8 1 1~8 35.34 14 8 1 1~8 48.33 15 7 2 2~7 29.08 16 6 1 1~6 42.93 21 3 1 1~3 30.93 22 8 1 1~8 51.13 25 7 1 1~7 76.17 29 4 1 1~4 38.24 33 3 1 1~3 15.07 35 6 4 4~6 10.00 36 7 6 6~7 7.19 37 7 6 6~7 5.70 38 4 3 3~4 14.34 39 8 1 1~8 45.39 43 9 1 1~9 110.89 表 4 基于研究材料的测量性状描述结果Table 4. Descriptions on measured qualities of maize under study性状编号
Characters
number最大值
Max最小值
Min极差
Range覆盖范围
Coverage
area平均值
Average
value变异系数
Coefficient of
variation/%17 47.6 21.7 25.9 21.7~47.6 37.3 14.02 18 37.5 15.4 22.1 15.4~37.5 27.6 15.11 19 24.5 1.8 22.7 1.8~24.5 9.1 53.42 20 30.5 9.7 20.8 9.7~30.5 21.3 20.67 23 112.0 42.0 70.0 42.0~112.0 90.1 14.21 24 12.1 5.9 6.2 5.9~12.1 10.0 11.65 26 208.6 24.4 184.2 24.4~208.6 115.6 31.42 27 433.7 137.4 296.3 137.4~433.7 290.9 19.55 28 0.6 0.2 0.4 0.2~0.6 0.4 18.93 30 21.7 11.1 10.6 11.1~21.7 17.3 11.37 31 5.6 3.5 2.1 3.5~5.6 4.8 8.46 32 18.5 11.8 6.7 11.8~18.5 14.8 9.63 2.2 玉米标准品种筛选结果
通过对100个试验材料进行田间种植和数据采集,并遵循《总论》中标准品种选择的原则进行筛选。标准品种筛选结果如表5所示,共筛选到玉米标准品种29个,包括1个糯质玉米自交系、8个普通玉米自交系、1个糯质玉米杂交种、6个甜玉米杂交种和13个普通玉米杂交种。标准品种可覆盖37个数量性状,示例159种表达状态,平均每个品种可示例5.5种性状表达状态。玉米品种AA128、毫发520可分别示例19个和18个性状的表达状态,S02可示例12个性状的表达状态,S01可示例11个性状的表达状态,DT1581可示例10个性状的表达状态,A001可示例8个性状的表达状态,DT12832、龙白玉999可示例7个性状的表达状态,龙白玉3号、瑞黑糯1号可示例6个性状的表达状态,瑞佳甜3号、砚白1号、宣农玉7号可示例5个性状的表达状态,DT5133、隆白玉109、万丰玉939可示例4个性状的表达状态,S03、TRWX-9、云甜玉12号、富好2号、瑞甜7号可示例3个性状的表达状态,腾丰玉198、田单60、西南玉99可示例2个性状的表达状态,双色年华、云甜玉11号、瑞喜731、云黄13号、雅玉2号可示例1种性状的表达状态。平均每个性状约有4.3个性状表达状态可被标准品种示例,其中,最多的性状有5个性状表达状态可被标准品种示例,如散粉期;最少的性状只有1个性状表达状态可被标准品种示例,如叶片绿色程度。
表 5 玉米标准品种筛选结果Table 5. Selected standard varieties of maize品种编号Varieties number 品种名称
Varieties name性状编号及表达状态代码
Characters number and code of expression status2 AA128 2(7),3(2),4(7),5(7),7(1),11(5),14(5),16(1),17(5),18(5),20(3),22(1),23(3),25(1),27(6),29(4),31(5),32(5),43(1) 4 A001 10(3),11(3),12(6),13(3),22(7),23(4),25(7),29(2) 5 S01 2(5),9(1),10(1),14(1),15(3),21(3),22(3),24(3),26(1),27(3),28(1) 6 S02 4(5),5(5),9(3),13(7),19(1),20(1),21(2),22(5),24(1),28(3),30(5),39(2) 7 S03 26(5),27(9),30(7) 9 DT12832 6(2),20(5),23(5),24(9),25(5),28(5),32(6) 13 TRWX-9 23(2),26(2),30(3) 14 DT5133 26(5),27(7),31(7),33(1) 18 DT1581 4(9),5(9),7(5),9(7),16(3),24(5),26(7),28(7),31(3),32(4) 28 龙白玉999 6(4),12(8),14(3),17(7),19(7),27(9),30(5) 30 龙白玉3号 19(5),24(9),26(9),31(7),32(7),33(2) 34 腾丰玉198 9(5),24(3) 37 隆白玉109 19(9),23(7),24(5),33(3) 42 云甜玉12号 23(3),28(1),36(7) 44 双色年华 6(6) 45 毫发520 4(3),5(3),13(1),17(3),18(3),19(3),23(1),24(1),26(1),27(1),28(3),30(3),31(5),32(5),36(6),37(6),38(3),39(6) 46 云甜玉11号 35(6) 47 瑞佳甜3号 26(3),27(3),35(4),37(7),38(4) 55 砚白1号 7(3),10(5),16(5),23(5),28(9) 58 宣农玉7号 13(5),26(7),27(7),32(3),39(4) 64 田单60 12(2),30(7) 75 西南玉99 26(5),43(5) 82 万丰玉939 15(7),18(7),25(3),28(5) 84 瑞喜731 43(7) 88 瑞黑糯1号 14(7),15(5),27(5),31(3),39(8),43(9) 89 云黄13号 12(4) 91 富好2号 10(7),20(7),28(7) 99 雅玉2号 43(3) 100 瑞甜7号 2(1),4(1),5(1) 第三列中括号前面的数字表示该标准品种可示例的性状编号,括号里的数字为该标准品种可示例的表达状态代码。
Datum in front of parentheses on 3rd column is code of agronomic trait; that within parentheses, code of expressive status.2.3 标准品种的聚类分析
以筛选得到的29个玉米标准品种形态性状数据为基础,利用NTSYSpc 2.10e统计软件对筛选的玉米标准品种进行聚类分析,分析结果如图1所示。由图可知,本试验筛选的29个玉米标准品种间的相似系数为
0.7841 ~0.9098 ,品种编号分别为28和34、44和45两对品种间的相似系数最高,均为0.9098 ,其余大多数品种间的相似系数在0.8051 ~0.8889 。以0.7841 为阈值时,可以将29个标准品种分为两个类群,第Ⅰ类群包含6个品种,约占21%,主要为甜质、糯质型杂交玉米品种;第Ⅱ类群包含23个品种,约占79%,主要包括1个糯质玉米自交系、8个普通玉米自交系和14个普通玉米杂交种。3. 讨论
3.1 玉米标准品种的筛选
《指南》[7]中提供的标准品种在某些性状覆盖的表达状态较多,在某些性状可示例的标准品种较少,示例分布不均衡,不能起到很好的示例矫正作用。品种的一致性和稳定性较好,就能很好地发挥参考作用[8]。当前《指南》中提供的标准品种主要以自交系为主,杂交中占很小的比例,而且在低纬高原生态区域生长适应性不好,所以性状表达差异比较明显,标准品种的作用未能得到很好的发挥。由于玉米自交系区域适应没有杂交种强,其性状表达较杂交种更容易受环境因素影响,所以本研究设计的材料中杂交种材料较自交系要多。最终筛选的29个标准品种中杂交种有20个品种,约占总数的70%,自交系有9个品种。从筛选结果来看,本试验筛选的29个标准品种在有些性状的两端表达状态未能发挥示例作用,在一定程度也说明本研究涉及材料的多样性不够。
3.2 玉米标准品种的示例作用分析
与《指南》[7]中提供的玉米标准品种相比,《指南》中涉及59个玉米标准品种,覆盖37个数量性状,示例164个表达状态,平均每个标准品种示例2.8个性状表达状态。2012年版本[21]中的标准品种共计55个,覆盖37个数量性状,示例159个表达状态,平均每个标准品种示例4.3个性状表达状态。本试验最终筛选了29个玉米品种作为标准品种,覆盖37个数量性状,示例159个表达状态,平均每个品种可示例5.5个性状表达状态。由此可以看出,29个标准品种尽管示例的性状表达状态数量没有比之前两个版本《指南》中标准品种示例的多,但平均每个品种可示例的性状表达状态数量较多,对整个测试工作降低成本、减少工作量的效果明显。通过比较两个测试指南版本中[7,21]的玉米标准品种,最新指南版本有59个玉米标准品种,之前版本中有55个玉米标准品种,但两者之间的差异品种只有8个。说明最新《指南》中的玉米标准品种大多数仍用十多年前筛选的标准品种,但由于全球气候环境的变化及品种自身对生态条件适应能力的减弱,我们有必要加强玉米标准品种筛选方面的研究,及时更换标准品种,真正使标准品种发挥作用。
3.3 玉米标准品种聚类分析
根据聚类结果可以知道筛选的玉米标准品种间的相似系数为
0.7841 ~0.9098 ,较前人[11−13]对标准品种研究结果要高,虽然筛选的标准品种性状覆盖范围大、示例性强,但品种间的相似性也比较高。以0.7841 为阈值时,可以将29个标准品种分为两个类群,第Ⅰ类群包含6个品种,主要为甜质、糯质型杂交玉米品种;第Ⅱ类群包含23个品种,主要为普通型玉米品种。从聚类结果来看,本试验筛选的29个标准品种间相似度较高,说明此次筛选的标准品种间的遗传背景差异不大。从另外一方面也说明本试验中使用的37个数量性状的品种区分力一般。但本试验筛选的29个玉米标准品种可示例159个表达状态,覆盖的性状表达状态还是相对比较高,标准品种筛选效果比较理想。4. 结论
通过本试验研究,从100个玉米试验材料中筛选到29个玉米品种可作为标准品种,品种间相似系数为
0.7841 ~0.9098 ,覆盖37个数量性状,示例159个表达状态,平均每个品种可示例约5.5个性状表达状态,品种的示例能力较《指南》中标准品种还强。该29个玉米品种可作为云南等低纬高原区域进行DUS测试的标准品种,为今后进一步使用和标准品种相关研究奠定基础和提供参考。 -
![]()
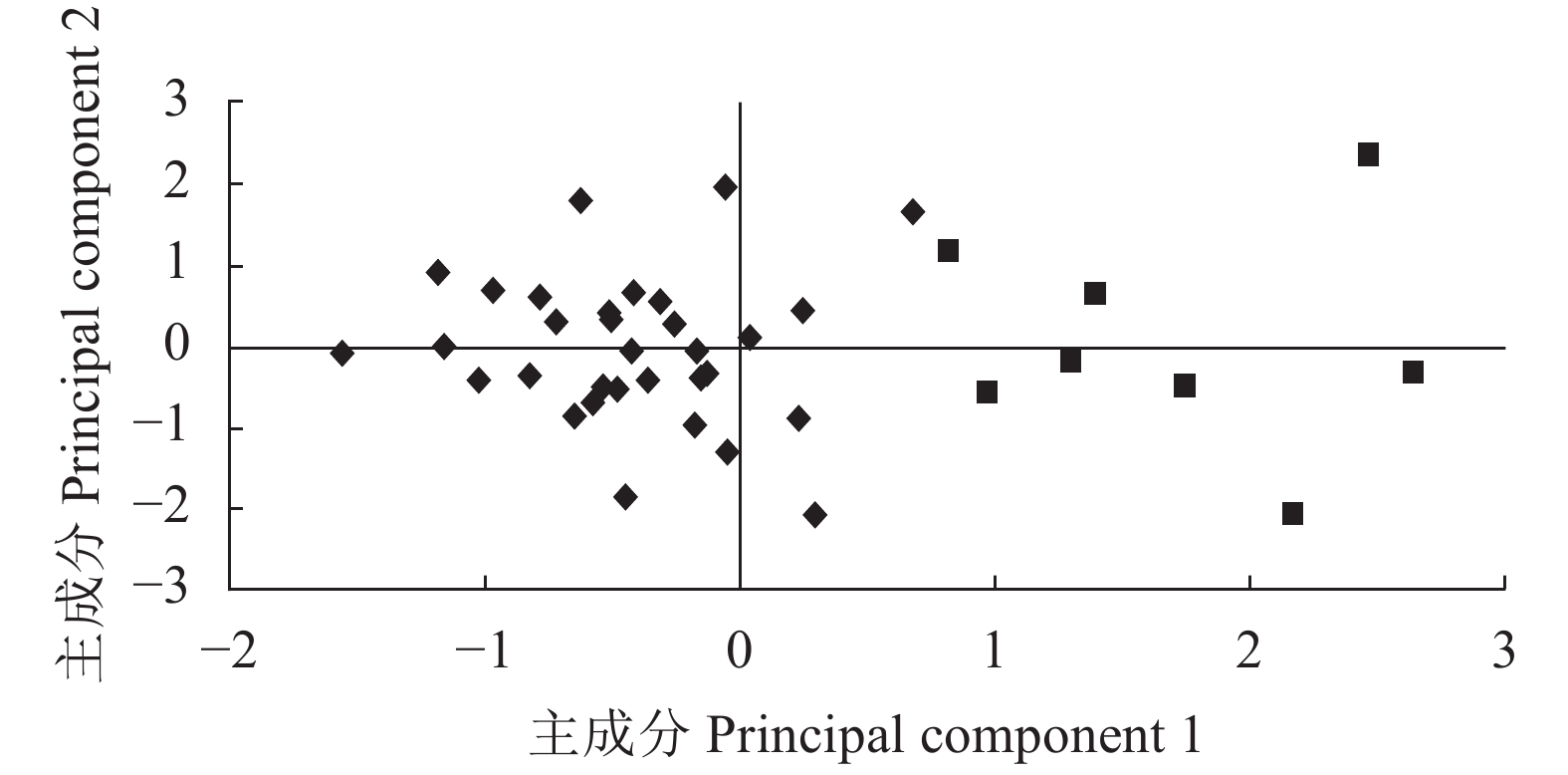
图 4 40份澳洲坚果种质资源花序表型性状主成分分布
Figure 4. Principal component distribution on phenotypic characteristics in inflorescence of 40 Macadamia germplasms
表 1 澳洲坚果种质资源名称、来源及类型
Table 1 Names, origins and types of Macadamia spp. in collection
编号
No.种质名称
Name of materials来源
Source种质类型
Germplasm types编号
No.种质名称
Name of materials来源
Source种质类型
Germplasm types1 HAES246 夏威夷 Hawaii M. integrifolia 21 B3-74 澳大利亚 Australia M. integrifolia 2 HAES294 夏威夷 Hawaii M. integrifolia 22 DAD 澳大利亚 Australia M. integrifolia 3 HAES333 夏威夷 Hawaii M. integrifolia 23 H2 澳大利亚 Australia M. integrifolia 4 HAES344 夏威夷 Hawaii M. integrifolia 24 NG18 澳大利亚 Australia M. integrifolia 5 HAES660 夏威夷 Hawaii M. integrifolia 25 O.C. 澳大利亚 Australia M. integrifolia 6 HAES695 夏威夷 Hawaii M. integrifolia/M. tetraphylla Hybrids 26 Own venture 澳大利亚 Australia M. integrifolia 7 HAES741 夏威夷 Hawaii M. integrifolia 27 Winks 澳大利亚 Australia M. integrifolia 8 HAES780 夏威夷 Hawaii M. integrifolia 28 Yonik 以色列 Israel M. integrifolia 9 HAES783 夏威夷 Hawaii M. integrifolia 29 南亚1号 Nanya 1 中国 China M. integrifolia 10 HAES788 夏威夷 Hawaii M. integrifolia 30 南亚2号 Nanya 2 中国 China M. integrifolia 11 HAES800 夏威夷 Hawaii M. integrifolia 31 南亚3号 Nanya 3 中国 China M. integrifolia 12 HAES812 夏威夷 Hawaii M. integrifolia 32 南亚12号 Nanya 12 中国 China M. integrifolia 13 HAES814 夏威夷 Hawaii M. integrifolia 33 南亚116号 Nanya 116 中国 China M. integrifolia 14 HAES826 夏威夷 Hawaii M. integrifolia 34 桂热1号 Guire 1 中国 China M. integrifolia 15 HAES842 夏威夷 Hawaii M. integrifolia 35 优株 Elite plant 24 中国 China M. integrifolia 16 HAES861 夏威夷 Hawaii M. integrifolia 36 优株 Elite plant 114 中国 China M. integrifolia 17 HAES900 夏威夷 Hawaii M. integrifolia/M. tetraphylla Hybrids 37 优株 Elite plant A 中国 China M. integrifolia 18 HAES915 夏威夷 Hawaii M. integrifolia 38 优株 Elite plant B 中国 China M. integrifolia 19 A4 澳大利亚 Australia M. integrifolia/M. tetraphylla Hybrids 39 优株 Elite plant D 中国 China M. integrifolia 20 A16 澳大利亚 Australia M. integrifolia/M. tetraphylla Hybrids 40 优株 Elite plant G 中国 China M. integrifolia  下载: 导出CSV
下载: 导出CSV
表 2 鉴定性状、鉴定部位及记载标准
Table 2 Properties, parts and standards for identifications
编号 No. 性状 Characters 鉴定部位 Identification parts 记载标准 Description of grading 1 小花颜色
Floweret color花序中开放小花的颜色
The color of opening floweret in the inflorescence1=乳白色 Creamy white,2=淡粉色 Light pink,3=粉红色 Pink 2 小花开放顺序
Floweret opening order花序中小花的开放顺序
Floweret opening order in the inflorescence1=自花序轴基部向顶端依次开放
Floweret opening sequentially from the base of inflorescence axis to the top,
2=由花序轴中部向两端依次开放
Floweret opening sequentially from the middle of inflorescence axis to the both ends,
3=自花序轴顶端向基部依次开放
Floweret opening sequentially from the top of inflorescence axis to the base3 批次开花
Flower in batches开花期有无分批次开花
Whether flower in batches or not1=有 Yes,2=无 Not 4 花序长度/cm
Inflorescence length花序主轴长度
the length of inflorescence axis5 小花长度/mm
Floweret length将要开放小花基部到顶部的长度
Length from base to top of floweret being to open6 小花数量/朵
Floweret number/Flower每个花序上着生小花的数量 Floweret number in the inflorescence
下载: 导出CSV
表 3 3 个描述性状多样性统计分析
Table 3 Statistical analysis on diversity based on 3 descriptive characteristics
性状
Characters评价
Evaluation种质份数
Germplasm copies/份百分比
Percentage/%小花颜色
Floweret color乳白色 Creamy white 36 90.00 淡粉色 Light pink 2 5.00 粉红色 Pink 2 5.00 小花开放顺序
Floweret opening order自花序轴基部向顶端依次开放
Floweret opening sequentially from the base of inflorescence axis to the top20 50.00 由花序轴中部向两端依次开放
Floweret opening sequentially from the middle of inflorescence axis to the both ends6 15.00 自花序轴顶端向基部依次开放
Floweret opening sequentially from the top of inflorescence axis to the base14 35.00 批次开花
Flower in batches有 Yes 9 22.50 无 Not 31 77.50
下载: 导出CSV
表 4 3 个数量性状多样性统计分析
Table 4 Statistical analysis on diversity based on 3 quantifiable characteristics
性状 Characters 平均值 Mean 最大值 Max 最小值 Min 标准差 SD 极差 R 变异系数 CV/% 花序长度 Inflorescence length/cm 18.06 29.0 11.7 4.75 17.3 26.30 小花长度 Floweret length/mm 8.02 9.4 6.6 0.69 2.8 8.60 小花数量 Floweret number/朵 Flower 177.0 277 130 37.10 147 20.96
下载: 导出CSV
表 5 主成分的特征向量、特征值、贡献率和累积贡献率
Table 5 Eigenvectors, latent roots, single and accumulative contributor ratios of principal components
性状
Characters主成分1
Principal component 1主成分2
Principal component 2主成分3
Principal component 3小花颜色 Floweret color 0.299 0.631 0.237 小花开放顺序 Floweret opening order −0.182 −0.548 0.654 批次开花 Flower in batches −0.161 0.604 0.602 花序长度 Inflorescence length 0.940 −0.061 −0.038 小花长度 Floweret length −0.327 0.494 −0.336 小花数量 Floweret number 0.936 0.031 0.076 特征值 Latent root 2.015 1.324 0.967 贡献率 Contributor ratio 33.577 22.064 16.112 累积贡献率
Accumulative contributor ratio33.577 55.640 71.752
下载: 导出CSV
-
[1] 张汉周, 王维, 杨为海, 等. 21份澳洲坚果种质开花结果物候期的变异分析 [J]. 热带作物学报, 2015, 36(11):2039−2043. DOI: 10.3969/j.issn.1000-2561.2015.11.019 ZHANG H Z, WANG W, YANG W H, et al. Variation analysis of blooming and fruiting phenophase of Macadamia spp. germplasm resources [J]. Chinese Journal of Tropical Crops, 2015, 36(11): 2039−2043.(in Chinese) DOI: 10.3969/j.issn.1000-2561.2015.11.019
[2] 万继锋, 杨为海, 曾辉, 等. 澳洲坚果种质资源叶片表型多样性分析及其数量分类研究 [J]. 热带作物学报, 2017, 38(6):990−997. DOI: 10.3969/j.issn.1000-2561.2017.06.002 WAN J F, YANG W H, ZENG H, et al. Leaf Character Diversity Analysis and Numerical Classification of Macadamia spp. Germplasm Resources [J]. Chinese Journal of Tropical Crops, 2017, 38(6): 990−997.(in Chinese) DOI: 10.3969/j.issn.1000-2561.2017.06.002
[3] 陆超忠,肖邦森,孙光明,等.澳洲坚果优质高效栽培技术[M]. 北京:中国农业出版社, 2000. [4] 云南省人民政府.2018年第八届国际澳洲坚果大会新闻发布会[EB/OL]. (2018-06-16). http://ynxwfb.yn.gov.cn/html/2018/wangqihuigu_0616/260.html. [5] 万继锋, 吴如健, 韦晓霞, 等. 橄榄种质资源叶片表型性状多样性分析 [J]. 热带作物学报, 2012, 33(6):975−980. DOI: 10.3969/j.issn.1000-2561.2012.06.002 WAN J F, WU R J, WEI X X, et al. Diversity analysis of the leaf phenotypic characters of Chinese olive germplasm resources [J]. Chinese Journal of Tropical Crops, 2012, 33(6): 975−980.(in Chinese) DOI: 10.3969/j.issn.1000-2561.2012.06.002
[6] ARADHYA M K, YEE L K, ZEE F T, et al. Genetic variability in Macadamia [J]. Genetic Resources and Crop Evolution, 1998, 45(1): 19−32. DOI: 10.1023/A:1008634103954
[7] 贺熙勇, 倪书邦, 陈国云, 等. 澳洲坚果种质资源形态性状的遗传多样性分析 [J]. 中国农学通报, 2010, 26(3):206−215. HE X Y, NI S B, CHEN G Y, et al. Morphological characteristics analysis on genetic diversity of Macadamia(Macadamia spp.)germplasm resources [J]. Chinese Agricultural Science Bulletin, 2010, 26(3): 206−215.(in Chinese)
[8] 杨为海, 王维, 曾辉, 等. 澳洲坚果不同种质果实数量性状的研究 [J]. 热带作物学报, 2011, 32(8):1434−1438. DOI: 10.3969/j.issn.1000-2561.2011.08.008 YANG W H, WANG W, ZENG H, et al. Fruit quantitative characters of various Macadamia germplasms [J]. Chinese Journal of Tropical Crops, 2011, 32(8): 1434−1438.(in Chinese) DOI: 10.3969/j.issn.1000-2561.2011.08.008
[9] 曾辉, 杨为海, 张明楷, 等. 澳洲坚果种质果实品质性状的多样性分析 [J]. 经济林研究, 2013, 31(1):70−74. ZENG H, YANG W H, ZHANG M K, et al. Diversity analysis of fruit quality characters in macadamia [J]. Nonwood Forest Research, 2013, 31(1): 70−74.(in Chinese)
[10] 宫丽丹, 倪书邦, 马静, 等. 不同来源澳洲坚果种质品质性状多样性分析 [J]. 中国南方果树, 2019, 48(1):32−35, 40. GONG L D, NI S B, MA J, et al. Diversity of quality characters of Macadamia germplasm from different sources [J]. South China Fruits, 2019, 48(1): 32−35, 40.(in Chinese)
[11] 陆超忠, 杜丽清. 澳洲坚果种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2008. [12] 邹明宏, 陆超忠, 梁李宏, 等. NY/T 1687-2009澳洲坚果种质资源鉴定技术规范[S]. 北京: 中国农业出版社. 2009. [13] 江锡兵, 龚榜初, 刘庆忠, 等. 中国板栗地方品种重要农艺性状的表型多样性 [J]. 园艺学报, 2014, 41(4):641−652. JIANG X B, GONG B C, LIU Q Z, et al. Phenotypic diversity of important agronomic traits of local cultivars of Chinese chestnut [J]. Acta Horticulturae Sinica, 2014, 41(4): 641−652.(in Chinese)
[14] 刘国彬, 曹均, 兰彦平, 等. 板栗总苞与坚果表型多样性及其相关关系研究 [J]. 经济林研究, 2014, 32(2):28−33. DOI: 10.3969/j.issn.1003-8981.2014.02.006 LIU G B, CAO J, LAN Y P, et al. Phenotypic diversity of involucres and nut in Castanea mollissima and their relationship [J]. Nonwood Forest Research, 2014, 32(2): 28−33.(in Chinese) DOI: 10.3969/j.issn.1003-8981.2014.02.006
[15] ZENELI G, KOLA H, DIDA M. Phenotypic variation in native walnut populations of Northern Albania [J]. Scientia Horticulturae, 2005, 105(1): 91−100. DOI: 10.1016/j.scienta.2004.11.003
[16] TRUEMAN S J. The reproductive biology of macadamia [J]. Scientia Horticulturae, 2013, 150: 354−359. DOI: 10.1016/j.scienta.2012.11.032
[17] 顾蕴洁, 王忠, 高煜珠. 环境因素对水稻颖花开闭影响的机理 [J]. 植物生理学报, 1993, 19(4):345−352. DOI: 10.3321/j.issn:1671-3877.1993.04.013 GU Y J, WANG Z, GAO Y Z. An investigation on the effectsof environmental factors on the opening and closure of florets in rice [J]. Acta Phytophysiologica Sinica, 1993, 19(4): 345−352.(in Chinese) DOI: 10.3321/j.issn:1671-3877.1993.04.013
[18] 李靖, 李庆军. 小桐子花的性别与开花时间的相互关系 [J]. 热带亚热带植物学报, 2009, 17(1):1−4. DOI: 10.3969/j.issn.1005-3395.2009.01.001 LI J, LI Q J. The correlationship between flowering sequence and floral gender in the inflorescence of Jatropha curcas L.(Euphorbiaceae) [J]. Journal of Tropical and Subtropical Botany, 2009, 17(1): 1−4.(in Chinese) DOI: 10.3969/j.issn.1005-3395.2009.01.001
[19] UPOV. Macadamia: M. integrifolia Maiden et Betche, M.tetraphylla L.A.S Johnson. Guidelines for the conduct of tests for distinctness, homogeneity and stability[M]. Geneva: International Union for the Protection of New Varieties of P1ant, 1987.
[20] 王鑫, 伍涛, 陶书田, 等. 梨花序不同序位坐果对果实发育及品质的影响 [J]. 西北植物学报, 2010, 30(9):1865−1870. Wang X, Wu T, Tao S T, et al. Pear Fruit Development and Quality at Different Fruit Positions of Inflorescence [J]. Acta Botanica Boreali-Occidentalia Sinica, 2010, 30(9): 1865−1870.(in Chinese)
-
期刊类型引用(12)
1. 潘浩男,潘贞珍,环秀菊,覃潇敏,郑树芳,陆宇明,康专苗,陶亮,王文林,覃振师. 澳洲坚果桂热1号人工授粉有效性及花粉直感研究. 果树学报. 2025(01): 151-161 .  百度学术
百度学术
2. 孔广红,陶丽,贺熙勇,倪书邦,陈丽兰,肖晓明. 不同授粉组合对澳洲坚果良种HAES863坐果及果实大小的影响. 中国果树. 2024(03): 93-97 . 百度学术
3. 孔广红,余文才,李玲,陶丽,贺熙勇. 云南西双版纳澳洲坚果开花与结实特性. 南方农业学报. 2024(04): 1089-1098 . 百度学术
4. 马静,李扬,陶亮,贺熙勇,龙青姨,宫丽丹. 澳洲坚果种质资源花序表型性状多样性分析. 云南农业大学学报(自然科学). 2024(04): 131-140 . 百度学术
5. 刘姚,张金云,吴世军. 广东省澳洲坚果种业发展现状与展望. 果树资源学报. 2023(01): 1-6 . 百度学术
6. 万继锋,卢智强,邹明宏,陈菁,宋喜梅,杨倩,董丽红,罗炼芳,曾辉. 澳洲坚果成熟期间果实内果皮颜色、果仁品质变化及最适采收期预测. 中国南方果树. 2023(06): 72-75+84 . 百度学术
7. 林玉虹,杨祥燕,蔡元保,潘如军,崔明勇,叶维雁,曾黎明,黄思婕,李冰. 68份澳洲坚果种质资源的表型分类研究. 南方农业学报. 2023(08): 2379-2387 . 百度学术
8. 万继锋,曾辉,邹明宏,陈菁,宋喜梅,杨倩,罗炼芳,陆超忠. 澳洲坚果种质资源果实表型多样性分析. 中国南方果树. 2022(06): 134-140 . 百度学术
9. 万继锋,曾辉,邹明宏,陈菁,罗炼芳,陆超忠. 八个澳洲坚果品种成年树抽梢、开花结果特性及栽培要点. 热带农业科学. 2021(03): 43-47 . 百度学术
10. 谭秋锦,韦媛荣,黄锡云,张涛,许鹏,宋海云,王文林,郑树芳. 10份澳洲坚果种质果实性状与营养成分分析. 果树学报. 2021(05): 672-680 . 百度学术
11. 万继锋,曾辉,邹明宏,陈菁,罗炼芳,陆超忠. 湛江地区澳洲坚果成年树抽梢生物学特性的观测分析. 中国南方果树. 2021(05): 65-70+75 . 百度学术
12. 项亚男. 澳洲坚果形态解剖学及产业研究进展. 南方农业. 2021(26): 151-152 . 百度学术
其他类型引用(0)




计量
- 文章访问数: 1370
- HTML全文浏览量: 443
- PDF下载量: 46
- 被引次数: 12
