

Growth and Development of Leptocybe invasa as Affected by Environmental Temperature
-
摘要:目的 致瘿性昆虫—桉树枝瘿姬小蜂(Leptocybe invasa)的生长发育、繁殖及分布主要受到温度的影响,而探明温度对桉树枝瘿姬小蜂生长发育的影响,可为该害虫的虫情监测和预测预报提供科学依据。方法 采用室内恒温接种、饲养的方法,测定了桉树枝瘿姬小蜂在19、22、25、28、31℃下各虫态的发育历期和发育速率,利用直线回归(图测法)、线性回归和 Logistic 回归方程分析了温度与各虫态发育速率的关系,并用直线回归方程求得各虫态发育起点温度和有效积温。结果 温度对桉树枝瘿姬小蜂卵至蛹各虫态、全世代的发育历期及成虫寿命均有显著影响,在19~31℃范围内,各虫态发育历期均随温度升高而逐渐缩短,发育速率与温度呈显著正相关。图测法直线回归方程更能准确拟合桉树枝瘿姬小蜂各虫态发育速率与温度之间的关系。桉树枝瘿姬小蜂卵、幼虫、蛹、成虫(存活)及全世代的发育起点温度分别为10.78、10.29、14.64、14.73和12.68℃,有效积温分别为86.96、588.24、92.59、104.17和712.34 d·℃,发育最适温度分别为25.68、25.65、24.58、26.42和23.84℃,发育适宜温区为12.21~35.48℃。2017—2019年,该害虫在江西赣南发生世代数理论值为4.27代,与实际观察结果一致。结论 温度是影响桉树枝瘿姬小蜂生长发育的重要因素,本研究结果可为预测该蜂的发育历期、发生区域、发生期和发生代数提供科学依据和技术支撑,尤其对该蜂成虫发生期预测预报和有效防控具有重要的指导意义。Abstract:Objective Correlations between the growth, reproduction, and distribution of Leptocybe invasa, an insect that causes galls in plants, and the surrounding temperature were studied to facilitate the monitoring and controlling the spread of the pest wasp.Method Duration and speed of each development state of the wasp were measured by inoculating and feeding the larvae indoor at 19, 22, 25, 28 and 31℃. Graphic, linear, and logistic regression analyses were applied to establish the correlations. Threshold and effective temperatures for the emergences of various stages of the insect were obtained using a linear regression model.Result Temperature was found to significantly affect not only at the stage from egg to pupa, but also the entire development and adult life of the insect. The increasing temperature hastened the insect development process. The graphic linear regression model seemed to more accurately describe their correlation than the other models. In Gannan, Jiangxi, the threshold emergence temperature of the eggs was found to be 10.78℃; that of the larva, 10.29℃; that of the pupa, 14.64℃; the adult survival threshold temperature, 14.73℃; and, the temperature for the development of an entire generation, 12.68℃. The accumulated effective temperature for the eggs was found to be 86.96℃; that for the larva, 588.24℃; that for the pupa, 92.59℃; that for the adult survival, 104.17℃; and, that for the development of an entire generation, 712.34 d·℃. And, the optimum temperature for the egg development, 25.68℃; for the larvae, 25.65℃; for the pupa, 24.58℃; for the adult survival, 26.42℃; and, for the development of an entire generation, 23.84℃ with a range between 12.21℃ and 35.48℃. The model estimated 4.27 generations of L. invasa had taken place between 2017 and 2019 which was in agreement with what was observed in the field.Conclusion Temperature was critical for the development of L. invasa. Understanding the correlation would aid in predicting the development, infestation time and area, and emerging generations of L. invasa for effective control of the pest.
-
中国受盐分影响的土地面积大,这些土地不利于番茄作物生长[1]。此外,作物苗期也是对盐分极为敏感的时期,苗的质量关系到番茄最终的产量和品质。因此,培育抗盐壮苗,是盐土栽培番茄的关键环节[2-3]。番茄是一种普遍种植的中等耐盐蔬菜,也是茄科作物研究中的模式作物[4]。近年来,国内外已开展了番茄种苗抗盐性方面的研究。研究发现,氮磷钾复合肥与生物发酵鸡粪一起配制成有机无机复混肥,处理番茄幼苗,均可以促进盐胁迫条件下幼苗的生长,增加叶片中脯氨酸、叶绿素含量和抗氧化酶活性,从而增加番茄产量[5-7]。另外,钙、硅等无机营养元素在增强番茄幼苗抗盐性方面也发挥重要的功能。例如,番茄分别经过0.5、5、25 mmol·L-1钙含量的营养液处理后,高钙含量处理的番茄株高、直径和生物量均较钙含量低的处理大,且根、茎、叶中钙含量较高,过氧化物酶和多酚氧化酶活性较强,表明钙参与了番茄植株抗氧化胁迫的调控[8]。同样,施0.83 mmol·L-1硅处理作物,在盐胁迫条件下,植株体内Na+含量低,而渗透调节物质如蔗糖和果糖的含量高,说明硅也有利于改善作物的耐盐性,能减轻盐分诱导的渗透胁迫和离子毒害[9]。综上,有机无机肥配施,以及钙、硅肥单施均能一定程度改善番茄幼苗的部分耐盐性状。因此,为了研发抗盐番茄幼苗的营养基质,进行优化组合不同水平的有机肥、三元复合肥和硅钙肥等与番茄抗盐性有关的肥料,并选用草炭:蛭石:珍珠岩(3:1:1) 为基质材料,比较研究这些不同配比的肥料营养基质分别在0(对照)、100 mmol·L-1(盐处理) NaCl下对“石头168”番茄幼苗生长生理特征的影响,以便筛选出能明显增强番茄幼苗抗盐性的较佳配方的营养基质。
1. 材料与方法
1.1 试验材料及栽培方法
2015年6月2日,在福州市埔垱村玻璃温室内进行番茄幼苗移栽试验。试验采用L9正交试验,其因子与水平主要参照中华人民共和国农业行业标准蔬菜育苗基质进行设计(表 1),试验用的塑料盆长、宽、高分别为14、16、12 cm。每盆种1株幼苗,每个处理3次重复。番茄品种属于“石头168”,取自福建省永泰县丰园蔬菜育苗有限公司,苗龄为29 d,株高均为9.9 cm左右。选择草炭:蛭石:珍珠岩(3:1:1) 为基质原料,分别加入不同水平的有机肥、复合肥和硅钙肥等基质营养。其中,有机肥中有机质含量为368.9 g·kg-1,全N含量为9.0 g·kg-1,全P含量为22.90 g·kg-1,全K含量为5.29 g·kg-1;复合肥中全N含量为15.0 g·kg-1,全P含量为150.0 g·kg-1,全K含量为150.0 g·kg-1;硅钙肥中SiO2≥250 g·kg-1,CaO≥320 g·kg-1。试验开始时,先用淡水浇灌23 d,待番茄苗恢复正常生长后,6月份25日开始分别进行0、100 mmol·L-1 NaCl溶液处理,浇灌量均为每盆300 mL,盐处理的番茄幼苗隔天浇灌1次盐水,其他时间同0 mmol·L-1 NaCl处理一样均用淡水,在每天上午的8:00~9:00进行浇水。7月18日,进行采样,以备检测番茄生长生理指标。
表 1 正交试验L9(33)各处理的施肥种类及用量Table 1. Orthogonal experiment design on fertilizations for pot culture处理 肥料用量/(g·kg-1) 有机肥 三元复合肥 硅钙肥 #1 4.05 2.70 0.54 #2 4.05 5.40 1.08 #3 4.05 8.10 2.16 #4 6.75 2.70 1.08 #5 6.75 5.40 2.16 #6 6.75 8.10 0.54 #7 12.15 2.70 2.16 #8 12.15 5.40 0.54 #9 12.15 8.10 1.08 1.2 指标的测定及处理
1.2.1 生长指标
定期使用钢卷尺测量番茄苗株高、叶长和叶宽,并利用游标卡尺分别测定茎粗。番茄幼苗采收后,一部分植株样品经洗净、吸干,称取鲜重后,在105℃烘箱杀青15 min,于80℃烘干至恒重,称取干重,并计算根冠比、壮苗指数和植株含水量。它们的计算公式分别为:
根冠比=根干重/地上部植株干重
植株含水量/%=(鲜重-干重)/鲜重×100%
壮苗指数=(茎粗/株高+根干物质量/地上部干物质量)×全株干重。
相对生长速度=(成株株高-定植株高)/试验天数
1.2.2 生理指标
新鲜的番茄幼苗叶片被采收后,及时测定其生理指标。其中,采用丙酮浸提法测定叶绿素含量,采用电导法测定叶片电解质的相对外渗率[10]。
1.3 数据处理
数据采用SAS 8.02软件进行Duncan′s法新复极差多重比较(P<0.05) 分析,并进行主成分分析。其他统计分析则采用Excel 2003处理。
2. 结果与分析
2.1 不同配方肥料在无土基质中对盐分条件下番茄幼苗植株形态参数的影响
盐分条件下,#8、#5、#7、#4、#9育苗营养基质的番茄幼苗形态指标值较优(表 2)。尤其是处理#8的番茄幼苗根系干重、地上部干重、全株干重及壮苗指数均显著优于其他育苗营养基质,增幅分别为21.95%~143.90%、1.81%~61.88%、3.27%~65.96%、21.11%~127.51%,这表明#8处理更有利于番茄幼苗植株生物量的累积,从而促进幼苗的茁壮生长。
表 2 不同配方肥料在无土基质中对盐分条件下番茄苗期植株形态指标的影响Table 2. Effect of fertilizations for soil-free culture on growth performance of S. lycopersicum seedlings under salt stress营养基质 叶长/cm 叶宽/cm 株高/cm 茎粗/mm 根鲜重/g 根干重/g 地上部鲜重/g 地上部干重/g 全株鲜重/g 全株干重/g 根冠比 壮苗指数 #1 8.1e 3.9cd 81.2e 5.76i 1.82h 0.61f 55.79e 9.58d 57.61e 10.19d 0.064f 0.723h #2 8.7bc 4.2ab 74.0i 7.06e 2.44e 0.73d 50.62g 7.54h 53.06g 8.27h 0.097a 0.880f #3 9.0a 4.1bc 81.8d 7.80b 1.91g 0.66e 48.43h 7.89g 50.34h 8.55g 0.078d 0.744g #4 8.9ab 4.3ab 87.7b 7.12d 2.80b 0.83b 54.75f 9.16e 57.54f 9.99e 0.090c 0.990c #5 8.5cd 3.9cd 85.0c 6.80g 2.52d 0.82b 66.48a 10.51b 69.00a 11.33b 0.078d 0.979d #6 8.1e 3.7d 78.5g 6.20h 1.47i 0.44g 41.89i 6.61i 43.36i 7.05i 0.067e 0.527i #7 8.4d 4.4a 88.5a 7.86a 2.05f 0.79c 62.91d 10.16c 64.96d 10.95c 0.077d 0.944e #8 8.7bc 3.9cd 78.0h 7.46c 2.59c 1.00a 63.43c 10.70a 66.02c 11.70a 0.093b 1.199a #9 8.8ab 3.9cd 79.0f 6.86f 4.19a 0.83b 64.68b 8.75f 68.87b 9.58f 0.095ab 0.997b 注:同列数据后不同小写字母表示不同育苗营养基质间显著差异性水平(P < 0.05),下表同。 2.2 不同肥料配方在无土基质中对番茄幼苗相对生长速度的影响
不同育苗营养基质的番茄幼苗在盐分处理前、后相对生长速度见表 3。经过盐分处理后,不同育苗营养基质的幼苗相对高生长速度均有明显下降。与对照相比,处理#1、#2、#3、#4、#5、#6、#7、#8、#9的降幅分别为43.81%、70.79%、46.15%、51.37%、55.79%、62.63%、50.66%、57.21%、60.44%(表 3)。其中,盐分处理前,处理#4番茄幼苗相对生长速度最大,其次是#5,最小的是#2。由此可知,不管是在有(或无)盐分条件下,处理#4的番茄幼苗高生长均表现较好。
表 3 不同配方肥料在无土基质中对番茄幼苗相对生长速度的影响Table 3. Effect of fertilizations for soil-free culture on plant height increasing rate of S. lycopersicum seedlings营养基质 相对生长速度/(cm·d-1) NaCl处理前 NaCl处理后 #1 2.10fA 1.18bB #2 2.02gA 0.59hB #3 2.21eA 1.19bB #4 2.55aA 1.24aB #5 2.42bA 1.07dB #6 1.98hA 0.74gB #7 2.29cA 1.13cB #8 2.29cA 0.98eB #9 2.25dA 0.89fB 注:同列数据后不同小写字母表示不同育苗营养基质间显著差异性水平(P < 0.05);同一行不同大写字母表示NaCl处理和未经NaCl处理间显著差异性水平(P < 0.05),下表同。 2.3 不同育苗基质对盐分条件下番茄幼苗植株各组织含水率的影响
盐分条件下,不同育苗营养基质对番茄幼苗各组织含水率具有明显的影响(表 4)。其中,处理#9番茄根系、地上植株部分和全株含水率较高,其次为#2幼苗的地上部和全株含水率,以及#4幼苗的根系含水率,而#8幼苗根系含水率及#4地上部植株、全株含水率则较低。
表 4 不同配方肥料在无土基质中对盐分条件下番茄苗期植株各组织含水率的影响Table 4. Effect of fertilizations for soil-free culture on moisture content in tissues of S.lycopersicum seedlings under salt stress营养基质 含水率/% 根系 地上部分 全株 #1 66.37g 82.83i 82.31h #2 70.08c 85.10b 84.41b #3 67.80e 83.71f 83.10f #4 70.20b 83.27g 82.65g #5 67.30f 84.19d 83.57d #6 69.93d 84.22c 83.74c #7 61.66h 83.85e 83.15e #8 61.58i 83.13h 82.29h #9 80.10a 86.47a 86.08a 2.4 不同配方肥料在无土基质中对番茄幼苗叶片叶绿素含量的影响
叶绿素是植物进行光合作用的主要色素,它在光合作用的光吸收中起核心作用,主要有叶绿素a(Chl.a)和b(Chl.b)。由表 5可知,不同育苗营养基质的番茄幼苗,经盐分处理后,其叶片的Chl.a、Chl.b和Chl.(a+b)含量均比对照明显降低,表明盐分处理对番茄幼苗叶片的光合作用具有抑制效应。然而,所有盐分处理的番茄叶片Chl.a/b比值却较对照均有明显增加(表 5)。这表明盐分处理虽然一方面会降低番茄幼苗叶片中Chl.a、Chl.b的绝对含量,但同时另一方面番茄幼苗自身又可以通过调高Chl.a/b比值,在一定程度上缓解盐分对番茄幼苗叶片吸收光能的不利影响。此外,在盐分条件下,处理#9番茄苗叶片Chl.a、Chl.b和Chl.(a+b)含量均相对较高,而#8幼苗叶片的Chl.a/b比值则较大(表 5),有利于番茄幼苗在盐分条件下维持较高的光合作用水平。
表 5 不同配方肥料在无土基质中对番茄幼苗叶绿素含量的影响Table 5. Effect of fertilizations for soil-free culture on chlorophyll content in leaves of S. lycopersicum seedlings under salt stress营养基质 NaCl/
(mmol·L-1)叶绿素a
/(mg·g-1FW)叶绿素b
/(mg·g-1FW)叶绿素(a+b)
/(mg·g-1FW)叶绿素a/b
/(mg·g-1FW)#1 0 1.037gA 0.562iA 1.599iA 1.845bB 100 1.001fB 0.503fB 1.504fB 1.990bA #2 0 1.119eA 0.723dA 1.842dA 1.548fB 100 0.797hB 0.450hB 1.246hB 1.772gA #3 0 1.055fA 0.567hA 1.622hA 1.861aA 100 0.683iB 0.367iB 1.050iB 1.860cA #4 0 1.120eA 0.621gA 1.741gA 1.804cB 100 0.905gB 0.488gB 1.393gB 1.853dA #5 0 1.153cA 0.670fA 1.822fA 1.722dB 100 1.089dB 0.594cB 1.683cB 1.834eA #6 0 1.198abA 1.066bA 2.264bA 1.123hB 100 1.103bB 0.765bB 1.868bB 1.442iA #7 0 1.128dA 0.702eA 1.830eA 1.607eB 100 1.071eB 0.588dB 1.659dB 1.823fA #8 0 1.196bA 1.122aA 2.318aA 1.065iB 100 1.098cB 0.547eB 1.644eB 2.009aA #9 0 1.199aA 1.046cA 2.244cA 1.146gB 100 1.178aB 0.803aB 1.981aB 1.467hA 注:同列同盐分(或无盐分)处理不同小写字母表示不同育苗营养基质间显著差异性水平(P < 0.05);同一育苗营养基质的同列不同大写字母表示NaCl处理和未经NaCl处理间显著差异性水平(P < 0.05)。 2.5 不同肥料配方在无土基质中对番茄幼苗叶片电解质相对外渗率的影响
不同育苗营养基质下番茄苗期叶片电解质相对外渗率对盐分的响应见图 1。与对照相比,处理#1、#2、#3、#4、#5、#6、#7、#8、#9的增幅分别为10.53%、39.75%、121.39%、0.89%、128.76%、13.55%、247.60%、14.75%、200.37%(图 1)。由此可知,在遭受盐渍环境时,营养基质处理#4、#6、#8的番茄幼苗叶片电解质相对外渗率的增幅相对较小,特别是#4处理的叶片电解质相对外渗率在有(或无)盐分条件下都很小(图 1),这有助于缓解盐分对细胞膜造成的损伤,使细胞膜透性增大变缓,细胞内的电解质外渗速度增幅降低,外液电导率增幅变小,从而有利于减缓番茄苗期的盐害。
![]() 图 1 不同配方肥料在无土基质中对番茄幼苗叶片电解质相对外渗率的影响注:同盐分(或无盐分)处理不同小写字母表示不同育苗营养基质间显著差异性水平(P<0.05);同一育苗营养基质下不同大写字母表示NaCl处理和未经NaCl处理间显著差异性水平(P < 0.05)。Figure 1. Effect of fertilizations for soil-free culture on electrolyte leakage of S.lycopersicum seedlings under salt stress
图 1 不同配方肥料在无土基质中对番茄幼苗叶片电解质相对外渗率的影响注:同盐分(或无盐分)处理不同小写字母表示不同育苗营养基质间显著差异性水平(P<0.05);同一育苗营养基质下不同大写字母表示NaCl处理和未经NaCl处理间显著差异性水平(P < 0.05)。Figure 1. Effect of fertilizations for soil-free culture on electrolyte leakage of S.lycopersicum seedlings under salt stress3. 讨论与结论
本研究与曹兵等[11]配制含有复合肥的常规番茄育苗基质和包膜尿素的控施肥育苗基质相比,盐分条件下含有硅钙肥的处理#1、#2、#3、#4、#5、#6、#7、#8、#9比非盐分条件下常规基质育苗的番茄壮苗指数分别增加了50.59%、64.38%、69.85%、68.97%、88.83%、16.00%、102.61%、133.13%、73.31%;除了#6外,其他处理比控施肥育苗的壮苗指数也相应增加了1.81%、11.13%、14.83%、14.23%、27.66%、36.97%、57.61%、17.17%。由此可见,尽管本试验处理番茄在遭受盐渍环境条件下,但培育的番茄幼苗质量仍优于非盐渍条件的控施肥育苗和常规育苗。此外,与番茄苗有机基质配方[12]相比,本试验所有处理在75 d时,番茄幼苗在株高、茎粗和生物量等重要生长指标方面具有明显的优势,这是因为有机肥的肥效释放速率相对较慢,不能迅速被番茄幼苗吸收,促进生长。而且,本试验处理#1~#9的番茄幼苗相对生长速度是有机无机复合番茄幼苗基质的3.19~5.74倍[13]。综上研究结果比较分析表明,无论是在盐分环境中,还是非盐分条件下,本试验掺入了硅钙肥的有机无机复合基质在番茄育苗方面具有明显的优势,尤其是#8、#4这2个番茄育苗基质,培育的幼苗在生长生理等方面均表现良好,为盐土栽培番茄后期的丰产奠定了良好的基础。这是因为硅钙肥中含有有效活菌数≥2亿·g-1,并同时富含活性硅、钙、镁、锌、铁、硼、铜、钛、钼等中微量元素,且特别添加了免深耕调理剂、蚯蚓蛋白酶和磷素活化剂等[14],因此更有利于和有机肥、化肥起协同效应,共同促进番茄幼苗茁壮生长。
-
![]()
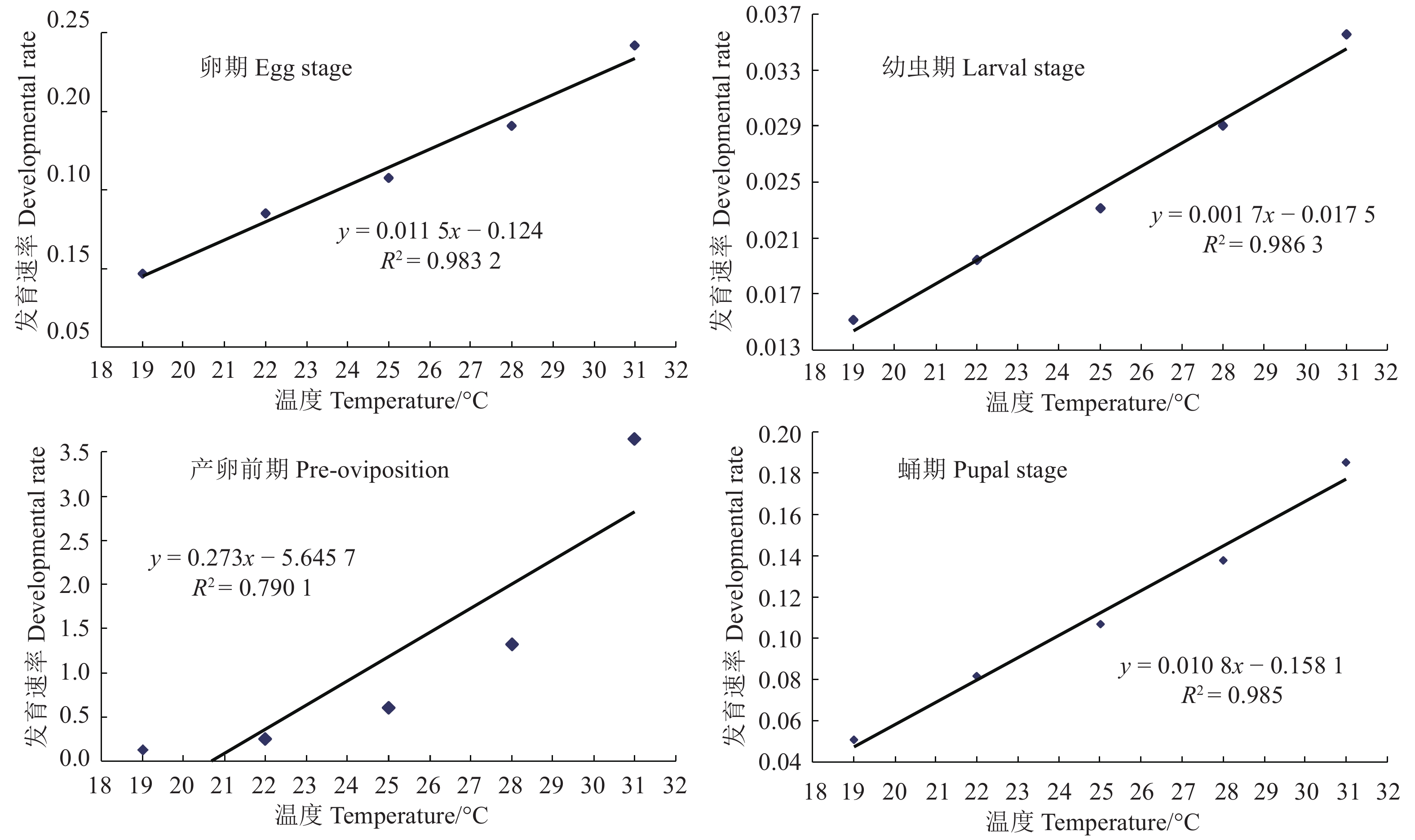
图 1 桉树枝瘿姬小蜂各虫态发育速率与温度的关系
Figure 1. Growth rate of L. invasa as affected by temperature
表 1 不同温度下桉树枝瘿姬小蜂各虫态的发育历期
Table 1 Durations of development stages of L. invasa under varied temperatures
虫态
Insect state发育历期 Developmental duration/d 19℃ 22℃ 25℃ 28℃ 31℃ 卵 Egg 10.33±1.56 aD 7.40±1.99 bD 6.33±1.73 cC 5.23±1.57 dC 4.33±1.24 eC 幼虫 Larva 66.13±4.58 aA 51.43±4.49 bA 43.27±4.65 cA 34.47±3.75 dA 28.17±3.21 eA 蛹 Pupa 19.63±2.41 aC 12.27±2.53 bC 9.37±1.77 cB 7.27±1.89 dB 5.40±1.30 eBC 产卵前期 Pre-oviposition 12.30±1.53 aD 5.13 ±2.34 bD 2.03 ±0.82 cD 1.05 ±0.52 dD 0.48 ±0.35 dD 成虫 Adult 23.97±3.23 aB 14.23±2.57 bB 10.33±1.63 cB 8.20±1.77 dB 6.27±1.57 eB 全世代 Generation 108.40±6.12 a 76.23±7.31 b 60.99±6.05 c 48.02±5.27 d 38.38±4.52 e 注:同行数据后不同小写字母表示同一阶段不同温度间差异显著,同列数据后不同大写字母表示同一温度不同世代间差异显著(SNK法, P<0.05);表中“成虫历期”指的是成虫寿命,世代历期=卵历期+幼虫历期+蛹历期+产卵前期,表2~4同。
Note: Different lowercase letters in the same row show significant difference, and different capital letters in the same column show significant difference(SNK, P<0.05);In the table, "adult duration" refers to adult life span;Generation duration = egg stage + larva stage + pupa stage + pre spawning stage. The same as Table 2–4. 下载: 导出CSV
下载: 导出CSV
表 2 桉树枝瘿姬小蜂各虫态发育速率及成虫寿命与温度的关系模型
Table 2 Regression models on growth rate and adult lifespan of L. invasa as affected by temperature
虫态
Insect state图测法 Graph method 线性回归 Linear regression Logistic回归 Logistic regression 模型
Model决定系数(R2)
Determination coefficient模型
Model决定系数(R2)
Determination coefficient模型
Model决定系数(R2)
Determination coefficient卵 Egg v=0.011 5t−0.124 0.983 2 v=0.012 1t−0.128 0 0.488 9 v=0.366 4/(1+e3.554 7−0.138 4t) 0.487 7 幼虫 Larva v=0.001 7t−0.017 5 0.986 3 v=0.001 7t−0.018 5 0.833 6 v=0.154 8/(1+e3.834 6−0.149 5t) 0.843 8 蛹 Pupa v=0.010 8t−0.158 1 0.985 0 v=0.011 8t−0.176 7 0.682 5 v=0.461 2/(1+e4.659 8−0.189 6t) 0.688 7 产卵前期 Pre-oviposition v=0.273t-5.645 7 0.790 1 v=0.273t-5.645 7 0.349 4 v=0.898 6/(1+e2.709 5−0.109 3t) 0.497 3 成虫 Adult v=0.009 6t−0.141 4 0.994 7 v=0.010 4t−0.156 9 0.733 9 v=0.305 9/(1+e4.792 7−0.181 4t) 0.736 0 全世代 Generation v=0.001 4t−0.017 8 0.992 4 v=0.001 4t−0.017 8 0.898 1 v=0.684 8/(1+e3.145 1−0.131 9t) 0.901 1
下载: 导出CSV
表 3 赣南桉树枝瘿姬小蜂各虫态的发育适温区范围
Table 3 Temperature ranges for optimal development at various stages of L. invasa in southern Jiangxi
虫态
Insect stateLogistic模型参数
Parameters of logistic model最适温Tmid
Optimum temperature/℃适温下限Tmin
Lower limit of temperature/℃适温上限Tmax
Upper limit of temperature/℃a b 卵 Egg 3.554 7 0.138 4 25.68 13.58 37.79 幼虫 Larva 3.834 6 0.149 5 25.65 13.85 37.45 蛹 Pupa 4.659 8 0.189 6 24.58 14.05 35.11 产卵前期 Pre-spawn 2.709 5 0.109 3 24.79 12.25 37.33 成虫 Adult 4.792 7 0.181 4 26.42 15.23 37.61 全世代 Generation 3.145 1 0.131 9 23.84 12.21 35.48
下载: 导出CSV
表 4 桉树枝瘿姬小蜂发育历期预测值与观测值的相关性分析
Table 4 Predicted and observed durations of L. invasa development stages
虫态
Insect state观测值 Observed value/d 预测值 Predicted value/d 相关系数(r)
Correlation coefficientP值
P Value19℃ 22℃ 25℃ 28℃ 31℃ 19℃ 22℃ 25℃ 28℃ 31℃ 卵 Egg 10.33 7.40 6.33 5.23 4.13 10.58 7.75 6.12 5.05 4.30 0.995 8 0.000 3 幼虫 Larva 66.13 51.43 43.27 34.47 28.17 67.57 50.25 40.00 33.22 28.41 0.994 3 0.000 5 蛹 Pupa 19.63 12.27 9.37 7.27 5.40 21.23 12.58 8.94 6.93 5.66 0.997 4 0.000 2 全世代 Generation 108.40 76.25 60.99 48.02 38.38 113.64 76.92 58.14 46.73 39.06 0.997 5 0.000 2
下载: 导出CSV
表 5 桉树枝瘿姬小蜂的LDT和SET (文献资料[10-13]及本试验)
Table 5 LDT and SET of L. invasa (data from literatures and this experiment)
地点
Position起点温度 LDT/℃ 有效积温 SET/(d·℃) 食料、温度
Food, Temperature卵
Egg幼虫
Larva蛹
Pupa成虫
Adult卵
Egg幼虫
Larva蛹
Pupa成虫
Adult福建福州
Fuzhou, Fujian 26.09°N8.65 6.57 13.94 14.83 101.80 807.16 105.43 114.53 巨桉8号, 21~36℃, 6个恒温[13]
E. grandis, 21~36℃, 6 constant temperatures江西赣州
Ganzhou, Jiangxi 25.85°N10.78 10.29 14.64 14.73 86.96 588.24 92.59 104.17 DH201-2, 19~31℃, 5个恒温(本研究结果)
DH201-2, 19~31℃, 5 constant temperatures [this experiment]广东广州
Guangzhou,Guangdong 23.08° N13.00 19.71 17.07 – 128.10 284.24 201.22 – 尾赤桉[12]
E. urophylla × E. camaldulensis广东广州
Guangzhou,Guangdong 23.08° N0.00 0.00 0.00 0.00 146.57 1228.01 161.67 205.17 DH201-2,室外,变温[11]
DH201-2, Outdoor, variable temperature广西南宁
Nanning, Guangxi 22.84°N10.25 10.79 749.43 53.42 20.5,27.5,28.6℃, 3个恒温[11]
20.5,27.5,28.6℃, 3 constant temperatures
下载: 导出CSV
-
[1] Honěk A. Geographical variation in thermal requirements for insect development [J]. European Journal of Entomology, 1996, 93: 303−312.
[2] JAROŠÍK V, HONĚK A, DIXON A F G. Developmental rate isomorphy in insects and mites [J]. The American Naturalist, 2002, 160(4): 497−510. DOI: 10.1086/342077
[3] BARTEKOVÁ A, PRASLIČKA J. The effect of ambient temperature on the development of cotton bollworm (Helicoverpa armigera Hübner, 1808) [J]. Plant Protection Science, 2010, 42(No. 4): 135−138.
[4] 陈元生, 罗致迪, 于海萍. 气象因子对赣南桉树枝瘿姬小蜂成虫发生量的影响 [J]. 中国植保导刊, 2016, 36(8):41−45. DOI: 10.3969/j.issn.1672-6820.2016.08.008 CHEN Y S, LUO Z D, YU H P. Effects of meteorological factors on adult population numbers of Leptocybe invasa in Southern Jiangxi Province [J]. China Plant Protection, 2016, 36(8): 41−45.(in Chinese) DOI: 10.3969/j.issn.1672-6820.2016.08.008
[5] 徐家雄, 任辉, 赵丹阳, 等. 桉树枝瘿姬小蜂种群发生规律与空间分布格局研究 [J]. 广东林业科技, 2008, 24(6):50−57. XU J X, REN H, ZHAO D Y, et al. Study on the population regularity and spatial distribution pattern of Leptocybe invasa at Eucalyptus [J]. Guangdong Forestry Science and Technology, 2008, 24(6): 50−57.(in Chinese)
[6] 陈元生, 涂小云, 罗益群. 赣南桉树枝瘿姬小蜂种群动态研究 [J]. 江苏农业科学, 2015, 43(11):178−180. CHEN Y S, TU X Y, LUO Y Q. Study on the population dynamics of Leptocybe invasa on Eucalyptus spp. in Southern Jiangxi [J]. Jiangsu Agricultural Sciences, 2015, 43(11): 178−180.(in Chinese)
[7] 陈元生, 廖忠明, 涂小云. 赣南桉树品种(系)对桉树枝瘿姬小蜂的抗性研究 [J]. 北方园艺, 2015(11):106−109. CHEN Y S, LIAO Z M, TU X Y. Study on resistance of different Eucalyptus varieties to leptocybeinvasain southern Jiangxi [J]. Northern Horticulture, 2015(11): 106−109.(in Chinese)
[8] 陈元生, 蔡润玉, 罗致迪, 等. 赣南桉树枝瘿姬小蜂种群物理监测技术研究 [J]. 现代农业科技, 2020(4):93−95, 97. CHEN Y S, CAI R Y, LUO Z D, et al. Study on population physical monitoring technology of Leptocybe invasa on Eucalyptus spp. in southern Jiangxi Province [J]. Modern Agricultural Science and Technology, 2020(4): 93−95, 97.(in Chinese)
[9] 陈元生, 金志芳, 涂小云. 桉树枝瘿姬小蜂化学防治药剂的筛选 [J]. 河南农业科学, 2015, 44(4):106−109. CHEN Y S, JIN Z F, TU X Y. Screening of chemical pesticides for control of Leptocybe invasa [J]. Journal of Henan Agricultural Sciences, 2015, 44(4): 106−109.(in Chinese)
[10] 吴耀军, 常明山, 李德伟, 等. 桉树枝瘿姬小蜂发育起点温度与有效积温的研究 [J]. 广东林业科技, 2013, 29(6):27−30. WU Y J, CHANG M S, LI D W, et al. Threshold and effective accumulative temperature for the development of Leptocybe invasa [J]. Guangdong Forestry Science and Technology, 2013, 29(6): 27−30.(in Chinese)
[11] 邱焕秀, 徐家雄, 林明生. 桉树枝瘿姬小蜂生物学特性和有效积温研究 [J]. 广东林业科技, 2011, 27(4):1−5. QIU H X, XU J X, LIN M S. Study on the biology and effective accumulated temperature by Leptocybe invasa at Eucalyptus [J]. Guangdong Forestry Science and Technology, 2011, 27(4): 1−5.(in Chinese)
[12] 朱方丽. 桉树枝瘿姬小蜂生物学及生态学研究[D]. 广州: 华南农业大学, 2012. ZHU F L. Study on biology and ecology of Leptocybe invasa at Eucalyptus[D]. Guangzhou: South China Agricultural University, 2012. (in Chinese)
[13] 陈汉章, 林秀琴, 陈顺立, 等. 桉树枝瘿姬小蜂发育起点温度和有效积温的测定 [J]. 长江大学学报(自科版), 2013, 10(29):1−6, 10, 8. CHEN H Z, LIN X Q, CHEN S L, et al. Measurement and application of developmental threshold temperature and effective cumulative temperature of Leptocybe invasa fisher et La salle [J]. Journal of Yangtze University(Natural Science Edition), 2013, 10(29): 1−6, 10, 8.(in Chinese)
[14] 牟吉元. 昆虫生态与农业害虫预测预报[M]. 北京: 中国农业科技出版社, 1997: 134-136. [15] 袁盛勇, 孔琼, 肖春, 等. 桔小实蝇各虫态发育历期及有效积温研究 [J]. 西南农业大学学报(自然科学版), 2005, 27(3):316−318. YUAN S Y, KONG Q, XIAO C, et al. Developmental threshold temperature and effective cumutive temperature of Bactrocera dorsalis hendel [J]. Journal of Southwest Agricultural University, 2005, 27(3): 316−318.(in Chinese)
[16] 陈元生, 罗致迪, 钟平华. 棉铃虫发育起点温度和有效积温的地理变异 [J]. 环境昆虫学报, 2017, 39(3):573−579. CHEN Y S, LUO Z D, ZHONG P H. Geographic variation indevelopment threshold temperature and sum of effective temperatures of the cotton bollworm, Helicoverpa armigera Hübner(Lepidoptera: Noctuidae) [J]. Journal of Environmental Entomology, 2017, 39(3): 573−579.(in Chinese)
[17] 宋墩福, 陈元生, 涂小云. 花绒寄甲赣南种群发育起点温度与有效积温研究 [J]. 中国植保导刊, 2015, 35(6):58−60, 44. DOI: 10.3969/j.issn.1672-6820.2015.06.015 SONG D F, CHEN Y S, TU X Y. Study on development threshold and sum of effective temperatures of Dastarcus helophoroides Gannan population [J]. China Plant Protection, 2015, 35(6): 58−60, 44.(in Chinese) DOI: 10.3969/j.issn.1672-6820.2015.06.015
[18] 潘雪红, 商显坤, 魏吉利, 等. 温度对蔗螟天敌等腹黑卵蜂生长发育的影响 [J]. 西南农业学报, 2019, 32(3):539−543. PAN X H, SHANG X K, WEI J L, et al. Effect of temperature on development of natural enemy Telenomus dignus [J]. Southwest China Journal of Agricultural Sciences, 2019, 32(3): 539−543.(in Chinese)
[19] 岳健, 何嘉, 张蓉, 等. 多异瓢虫的发育与温度的关系 [J]. 昆虫知识, 2009, 46(4):605−609. YUE J, HE J, ZHANG R, et al. Relationship between the temperature and the development of Hippodamia variegata [J]. Chinese Bulletin of Entomology, 2009, 46(4): 605−609.(in Chinese)
[20] 商显坤, 黄诚华, 魏吉利, 等. 温度对突背蔗犀金龟蛹发育速率的影响 [J]. 植物保护, 2017, 43(6):118−122. DOI: 10.3969/j.issn.0529-1542.2017.06.019 SHANG X K, HUANG C H, WEI J L, et al. Effects of temperature on the developmental rate of Alissonotum impressicolle pupae [J]. Plant Protection, 2017, 43(6): 118−122.(in Chinese) DOI: 10.3969/j.issn.0529-1542.2017.06.019
[21] 汤行昊. 桉树枝瘿姬小蜂生物学、生态学特性及防治试验[D]. 福州: 福建农林大学, 2013. Tang X H. Study on Biology, Ecology and control of Leptocybe invasa Fisher&La salle[D]. Fuzhou: Fujian Agriculture and Forestry University, 2013. (in Chinese)
[22] 时培建, 池本孝哉, 戈峰. 温度与昆虫生长发育关系模型的发展与应用 [J]. 应用昆虫学报, 2011, 48(5):1149−1160. DOI: 10.7679/j.issn.2095-1353.2011.189 SHI P J, IKEMOTO T K, GE F. Development and application of models for describing the effects of temperature on insects' growth and development [J]. Chinese Journal of Applied Entomology, 2011, 48(5): 1149−1160.(in Chinese) DOI: 10.7679/j.issn.2095-1353.2011.189
[23] 何莉梅, 葛世帅, 陈玉超, 等. 草地贪夜蛾的发育起点温度、有效积温和发育历期预测模型 [J]. 植物保护, 2019, 45(5):18−26. HE L M, GE S S, CHEN Y C, et al. The developmental threshold temperature, effective accumulated temperature and prediction model of developmental duration of fall armyworm, Spodoptera frugiperda [J]. Plant Protection, 2019, 45(5): 18−26.(in Chinese)
[24] LAMB R J. Developmental rate of Acyrthosiphon pisum (Homoptera: Aphididae) at low temperatures: implications for estimating rate parameters for insects [J]. Environmental Entomology, 1992, 21(1): 10−19. DOI: 10.1093/ee/21.1.10

计量
- 文章访问数: 1224
- HTML全文浏览量: 424
- PDF下载量: 19
