

Effect of Endogenous Hormones and Expressions of Relevant Genes on Tuber Growth of Bikeqi Yam
-
摘要:目的 分析毕克齐山药中内源激素含量与淀粉含量、可溶性总糖含量和还原糖含量的关系,以及与其相关合成基因表达量的相关性,探究毕克齐山药块茎膨大的机理。方法 以毕克齐山药5个不同生长期块茎为材料,采用酶联免疫吸附法分别测定脱落酸(ABA)、赤霉素(GA3)、生长素(IAA)、茉莉酸(JA)、玉米素(ZR)、异戊烯基腺苷(IPA)等6种内源激素含量,采用高效液相色谱仪测定方法测定水杨酸(SA)含量。结果 IAA、ZR、ABA、JA和SA含量与山药块茎形态指标正相关;GA3、IPA含量与形态指标负相关;IAA含量与山药块茎周长和块茎直径显著正相关;GA3含量与块茎长度显著负相关;与IAA相关的基因与内源激素IAA含量显著负相关。结论 内源激素IAA、ZR、ABA、JA和SA促进山药块茎膨大;GA3、IPA抑制山药块茎生长;IAA促进山药增粗;GA3抑制其伸长生长;与IAA相关的基因的下调表达对IAA的合成有促进作用,即正调控IAA含量。Abstract:Objective Effects of endogenous hormones and expressions of genes related to the hormone syntheses on the morphology and physiology of bikequ yam (Dioscorea Opposita) tubers during expanding stage were investigated. Correlation between the indicators was established to provide guidelines for upgrading the yield and quality of the yam farming.Method During expansion stage of the tubers, contents of starch, reducing sugar, and soluble total sugar were determined by chemical analysis, those of endogenous ABA, GA3, IAA, JA, ZR, and IPA by enzyme-linked immunosorbent assay, and that of salicylic acid (SA) by HPLC. Correlations between these indicators were tested by regression analysis. The gene expression levels of endogenous hormone related genes were obtained by transcriptome sequencing.Result The contents of IAA, ZR, ABA, JA, and SA correlated positively with the morphological indices on the tuber, while those of GA3 and IPA correlated negatively. IAA correlated with the tuber perimeter and diameter. GA3 correlated inversely with the length of tuber. The IAA-related gene was found to negatively correlate with IAA in the tubers.Conclusion Among the endogenous hormones, IAA, ZR, ABA, JA, and SA promoted the tuber enlargement; GA3 and IPA inhibited the growth; IAA enhanced the girth increase; and, GA3 retarded the elongation. The downregulation of IAA-related gene increased the hormone synthesis. In other words, the gene could regulate the IAA production in the yams, and in turn, affected the tuber growth.
-
0. 引言
【研究意义】山药(Dioscorea Opposita)块茎含有丰富的淀粉、蛋白质和多糖[1],其形态形成是复杂的生理生化过程,受相关基因和激素调控[2-3]。现已知的几种植物激素有脱落酸(abscisic acid,ABA)、生长素(indoleacetic acid,IAA)、茉莉酸(jasmonic acid,JA)、水杨酸(salicylic acid,SA)、赤霉素类(gibberellin,GA)以及细胞分裂素(cytokinin,CTK),其中细胞分裂素还包括玉米素(trans-Zeatin-riboside,ZR)和异戊烯基腺苷(isopentenyl adenosine,IPA)等[4]。探究内源激素及其相关基因表达量对山药块茎膨大的影响,可以为进一步从分子水平揭示山药膨大的机理提供理论依据。【前人研究进展】梁任繁[5]等认为山药块茎中积累的可溶性糖是影响块茎形成和发育的重要因素之一。周芸伊[6]研究发现在山药块茎膨大过程中激素调控发挥重要作用。周芸伊等[7]和甘立军等[8]在关于植物地下块茎的研究中发现,赤霉素被认为是与块茎形成关系最紧密的激素,块茎形成与内源赤霉素浓度密切相关。汪雷等[9]在对半夏生理特性的相关研究中发现,茉莉酸对植物根的生长,球茎的形成等生长发育过程有明显调控作用。Roumeliotis E等[10]和Zhang Z[11]的研究认为生长素对马铃薯块茎形成和发育有促进作用。细胞分裂素在植物根和茎的形态建成中也起到重要的调控作用[12]。说明内源激素对地下块根、块茎类植物的生长发育和膨大起到了显著影响。【本研究切入点】山药生长发育过程中淀粉含量、糖含量及内源激素含量与块茎膨大关系的研究尚少。【拟解决的关键问题】本研究通过毕克齐山药淀粉、可溶性总糖、还原糖和内源激素含量的变化,以及与内源激素合成相关基因的表达量的变化,探究这些因素对山药块茎膨大的影响,为进一步从分子水平的研究提供理论依据,同时也为山药的规模化生产及优质高产提供理论指导。
1. 材料与方法
1.1 试验材料
试验材料于2019年4月种植,同年10月收获。试验材料选择内蒙古当地栽培品种毕克齐山药(D.opposite Thunb.)。供试材料种植于呼和浩特市内蒙古农业大学种质资源圃,北纬40°29′28.01″,东经111°47′07.69″,土壤质地为砂壤土,地势平坦,灌溉方便。试验采用双行种植,大行距70 cm,小行距(沟内2列山药的行距)50 cm,株距30 cm,播种深度15~20 cm。
1.2 试验方法
山药块茎生长发育时期通常划分为3个时期,即:块茎膨大前期(种植后105~120 d)、块茎膨大中期(种植后120~150 d)、块茎膨大后期(种植后150~180 d)。本研究为了更详细完整地了解山药块茎膨大过程中的各形态指标、生理指标及内源激素含量的变化,在山药种植后105 d开始取样,每隔15 d取一次样,5次取样时间分别是种植后105 、120 、135 、150 、165 d。分别取5个取样点的山药块茎,每个取样点分别取3株作为生物学重复。分别测定形态指标、淀粉含量、糖含量、内源激素含量等。
取样时,将挖出的块茎放入自封袋中,迅速放入装有生物冰袋的取样箱中,以保持新鲜。带回实验室后,清水洗净、擦干,测量其形态指标;然后分别称取19 g新鲜样品进行淀粉、糖含量等指标的测定。每个品种分别称取2 g块茎,分别称取3次重复样,用锡纸包好后立即放入液氮中速冻,再放入−80 ℃冰箱中保存,以备后续内源激素的测定以及转录组测序。
淀粉、糖含量采用常规方法测定;样品中内源激素ABA、GA3、IAA、JA和ZR含量的测定方法同文献[12];内源激素SA含量采用高效液相色谱方法(HPLC)测定,提取方法参照Pan X[13]的方法并改进。
内蒙古农业大学山药种质资源课题组对毕克齐山药进行了转录组测序(2017年委托广州基迪奥生物科技有限公司测序),本研究结合已获得的转录组数据,筛选出与内源激素相关的差异表达基因,并分析了内源激素与其相关基因表达量的关系。
1.3 数据处理与分析
试验数据用Microsoft Excel 2016软件计算毕克齐山药的各形态指标、生理指标及内源激素含量3次重复的平均值并制图;运用SPSS 25.0对各指标进行方差分析(ANOVA)以Duncan法检测差异显著性,差异水平P<0.05;对形态指标、生理指标及内源激素进行相关性分析。
2. 结果与分析
2.1 毕克齐山药膨大过程淀粉含量等生理指标的变化趋势
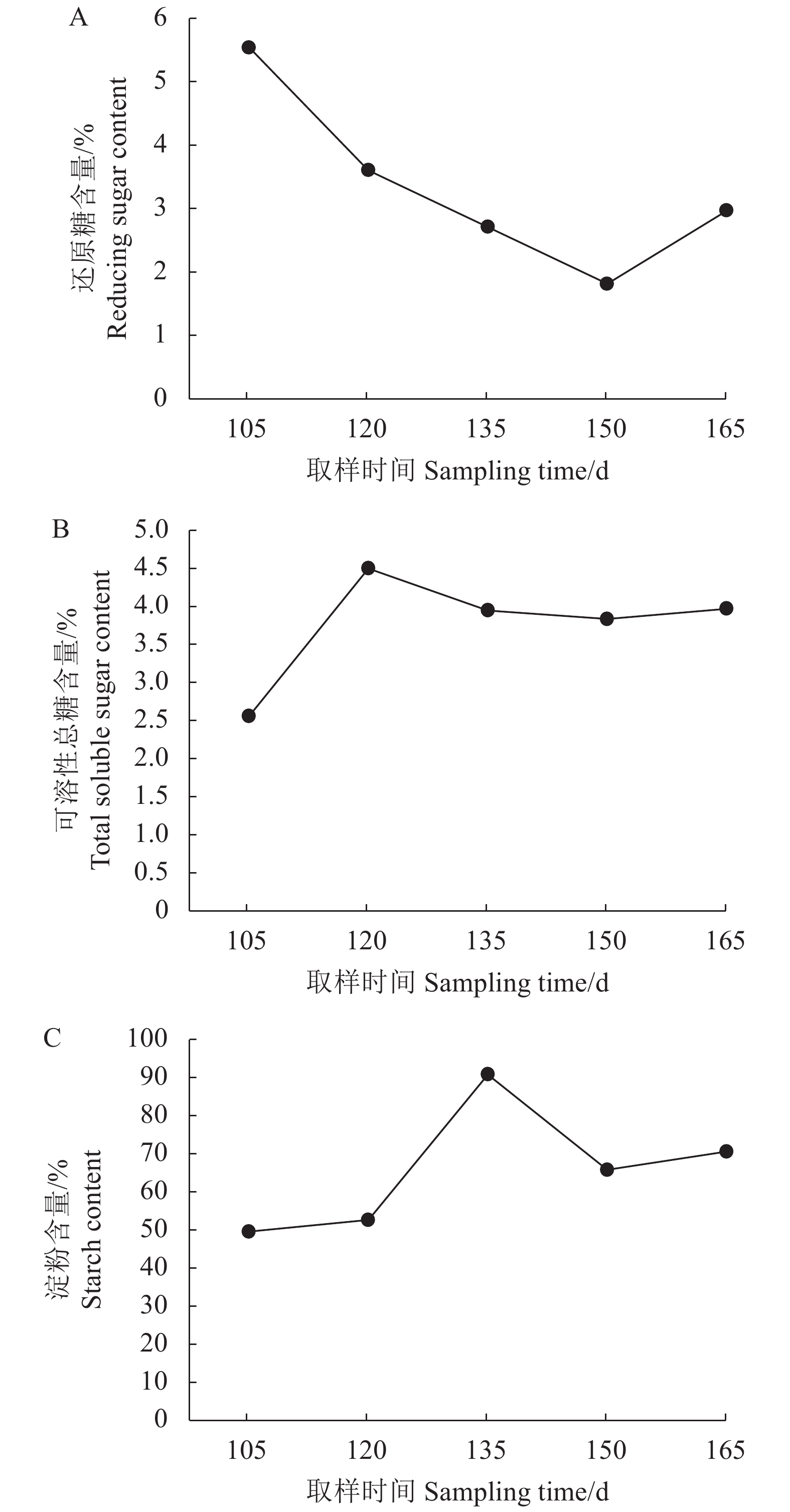
在毕克齐山药块茎的膨大过程中,对淀粉含量、还原糖含量和可溶性总糖含量进行平均值处理,得到各生理指标的变化趋势图(图1)。
![]() 图 1 生长发育过程各个生理指标的变化注:A-生长发育过程还原糖含量的变化;B-生长发育过程可溶性总糖含量的变化;C-生长发育过程淀粉含量的变化。Figure 1. Physiological changes on Bikeqi yam during tuber developmentNote: A-on reducing sugars; B-on soluble sugars; C-on starch.
图 1 生长发育过程各个生理指标的变化注:A-生长发育过程还原糖含量的变化;B-生长发育过程可溶性总糖含量的变化;C-生长发育过程淀粉含量的变化。Figure 1. Physiological changes on Bikeqi yam during tuber developmentNote: A-on reducing sugars; B-on soluble sugars; C-on starch.由图1可知,毕克齐山药的还原糖含量在块茎膨大过程中总体表现为下降趋势,即块茎膨大前期和块茎膨大中期呈下降趋势,块茎膨大后期小幅度上升,其中在第150 d时低于其他4个取样时间点。可溶性总糖含量先升后降趋势,种植120 d可溶性总糖含量达到高点之后下降,135~165d维持一定高度。淀粉含量总体表现为上升变化,在种植135 d达到最大值,之后略在小幅下降,150~165又开始小幅攀升。
2.2 内源激素含量的变化
由表1可知,IAA含量总体上表现为上升趋势,在第135 d时显著高于第105 d和第120 d这2个取样点(P<0.05),第165 d的IAA含量高于第150 d。ZR含量在块茎膨大过程中表现出先上升后下降的变化,并且差异显著,其中在第150 d时ZR含量最高,高于其他4个取样时间点。ABA含量总体呈上升变化,在第135 d最低,显著低于其他4个取样时间点(P<0.05),在第165 d时达到最大值,高于之前所有取样时间点。JA总体表现为上升趋势,第165 d时JA含量显著高于其他4个取样时间点(P<0.05)。GA3含量呈下降变化,最大值在第105 d并且显著高于之后4个取样时间点(P<0.05)。IPA含量在块茎膨大前期上升,在块茎膨大中期下降,在块茎膨大后期变化不明显,含量变化较小,未达到统计学显著水平。SA含量总体呈上升趋势,在第165 d时达到最大值,高于其他4个取样时间点。
表 1 毕克齐山药在块茎膨大过程内源激素含量变化Table 1. Changes on endogenous hormones during tuber enlargement of Bikeqi yam[单位: ng·g−1(FW)] 项目 Item 105 d 120 d 135 d 150 d 165 d 生长素 IAA 24.697±1.82 b 38.542±1.76 b 46.454±3.46 a 37.793±2.41 b 48.012±2.68 a 玉米素 ZR 5.295±0.23 b 6.97±0.39b 8.129±0.45 ab 9.39±0.72 a 6.751±0.36 b 脱落酸 ABA 33.274±1.93 bc 33.564±0.03 bc 28.402±1.70 c 40.707±2.11 b 81.939±1.88 a 茉莉酸 JA 11.976±0.43 b 12.678±0.39 b 15.806±1.02 ab 13.541±0.92 b 21.276±1.46 a 赤霉素 GA3 5.484±0.44 a 2.538±0.09 c 4.072±0.36 bc 3.209±0.20 bc 3.69±0.26 bc 异戊烯基腺苷 IPA 4.103±0.20 b 8.005±0.30 a 5.943±0.37 ab 4.202±0.21 b 4.419±0.27 b 水杨酸 SA 52.308±30.38 b 88.763±63.26 ab 19.226±10.83 b 81±37.48 ab 234.867±111.44 a 注:同行不同小写字母表示在0.05水平上差异显著。
Note: Data with different lowercase letters indicate significant differences at 0.05 level.2.3 内源激素含量与形态、生理指标的相关性
对毕克齐山药在整个生长发育过程中内源激素含量、形态指标和生理指标的平均值进行相关性分析,结果见表2。毕克齐山药的淀粉含量与形态指标呈正相关,还原糖含量与形态指标呈负相关,但均未达到统计学显著水平;可溶性总糖含量与各形态指标呈负相关,其中与块茎长度呈显著负相关(P<0.05),相关系数为-0.94;IAA与形态指标呈正相关,其中与块茎周长和块茎直径呈显著正相关(P<0.05),相关系数分别为0.906和0.904;ZR、ABA、JA和SA与形态指标呈正相关,但均未达到统计学显著水平;GA3与形态指标呈负相关,其中与块茎长度呈显著负相关(P<0.05),相关系数为-0.892;IPA与块茎长度呈正相关,与其余形态指标呈负相关关系,但差异均不显著。IAA与可溶性总糖含量呈显著负相关(P<0.05),相关系数为−0.883;ZR与还原糖含量呈显著负相关(P<0.05),相关系数为−0.932。
表 2 毕克齐山药内源激素与形态、生理指标相关性Table 2. Correlation between endogenous hormones and morphology and physiology of Bikeqi yam tubers项目
Item块茎长度
Length块茎周长
Circumference块茎直径
Diameter块茎鲜重
Fresh
weight块茎干重
Dry
weight淀粉含量
Starch
content还原糖含量
Reducing sugar
content可溶性总糖含量
Total soluble
sugar contentIAA ZR ABA JA GA3 IPA SA 块茎长度
Length0.756 0.757 0.674 0.679 0.375 −0.78 −0.940* 0.841 0.562 0.431 0.531 −0.892* 0.435 0.494 块茎周长
Circumference1.000** 0.922* 0.981** 0.719 −0.823 −0.839 0.906* 0.589 0.64 0.833 −0.425 −0.161 0.553 块茎直径
Diameter0.924* 0.982** 0.715 −0.822 −0.838 0.904* 0.588 0.643 0.833 −0.425 −0.162 0.557 块茎鲜重
Fresh weight0.972** 0.489 −0.84 −0.75 0.692 0.657 0.665 0.694 −0.43 −0.362 0.591 块茎干重
Dry weight0.646 −0.817 −0.771 0.807 0.602 0.677 0.804 −0.36 −0.321 0.579 淀粉含量
Starch content−0.617 −0.611 0.767 0.542 0.072 0.524 −0.061 −0.04 −0.082 还原糖含量
Reducing sugar content0.927* −0.732 −0.932* −0.21 −0.375 0.673 −0.034 −0.170 可溶性总糖含量
Total soluble sugar content−0.883* −0.773 −0.289 −0.494 0.805 −0.315 −0.293 IAA 0.483 0.479 0.779 −0.533 0.227 0.433 ZR −0.116 0.043 −0.575 0.022 −0.159 ABA 0.871 −0.118 −0.378 0.975** JA −0.107 −0.242 0.787 GA3 −0.612 −0.252 IPA −0.222 注:*在0.05水平,相关性显著。**在0.01水平,相关性极显著。表5、6同。
Note:*Significant correlation (P<0.05); **Extremely significant correlation (P<0.01). Same for Tables 5 and 6.2.4 内源激素合成相关基因表达量
内蒙古农业大学山药种质资源课题组对毕克齐山药进行了转录组测序,本研究从毕克齐山药转录组测序结果中筛选出与山药内源激素合成的相关基因共18个,其中与IAA合成相关的基因9个,与GA3合成相关的基因9个(表3)。
表 3 与内源激素合成相关基因的基因编号、基因代号和基因注释信息Table 3. Number, code, and information on genes associated with endogenous hormone synthesis in Bikeqi yam tubers项目
Item基因
ID Gene ID基因代号
Gene code基因注释信息
Gene annotation information与IAA合成相关基因 Genes related to IAA synthesis Unigene0016848 IAA 1 生长素响应蛋白基因 Auxin response protein gene Unigene0021936 IAA 2 生长素响应蛋白基因 Auxin response protein gene Unigene0023040 IAA 3 生长素响应蛋白基因 Auxin response protein gene Unigene0025388 IAA 4 IAA-氨基酸水解酶 ILR1基因 IAA-amino acid hydrolase ILR1 gene Unigene0027417 IAA 5 IAA-氨基酸水解酶 ILR1基因 IAA-amino acid hydrolase ILR1 gene Unigene0027794 IAA 6 生长素响应蛋白基因 Auxin response protein gene Unigene0028701 IAA 7 生长素响应蛋白基因 Auxin response protein gene Unigene0028916 IAA 8 生长素响应蛋白基因 Auxin response protein gene Unigene0030426 IAA 9 生长素响应蛋白基因 Auxin response protein gene 与 GA3合成相关基因 Genes related to GA3 synthesis Unigene0002430 GA3 1 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0019924 GA3 2 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0021502 GA3 3 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0021503 GA3 4 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0027812 GA3 5 赤霉素20氧化酶 Gibberellin 2-β-dioxygenase Unigene0023907 GA3 6 赤霉素调节蛋白基因 Gibberellin regulatory protein gene Unigene0024440 GA3 7 赤霉素调节蛋白基因 Gibberellin regulatory protein gene Unigene0026445 GA3 8 赤霉素调节蛋白基因 Gibberellin regulatory protein gene Unigene0020765 GA3 9 赤霉素调节蛋白基因 Gibberellin regulatory protein gene 由表4可知,毕克齐山药中IAA合成相关的基因表达量从总体上看,除少数例外,大部分rpkm值(Reads Per Kilobase per Million mapped reads)在块茎膨大过程中表现为下降趋势。其中IAA 3的表达量,在种植后105~135 d的下降幅度最大,rpkm值由213.97下降至119.15;IAA 4和IAA 7的表达量变化相似,二者均在块茎膨大前期和中期有一个上调表达,之后的表达量为下降变化。
表 4 与内源激素合成相关基因的rpkm平均值Table 4. Average rpkm of genes associated with endogenous hormone synthesis项目
Item基因ID
Gene ID基因代号
Gene code105 d 120 d 135 d 150 d 165 d 与IAA合成相关基因
Genes related to IAA synthesisUnigene0016848 IAA 1 95.82±30.63 65.29±44.90 64.25±22.21 26.73±12.83 26.68±10.96 Unigene0021936 IAA 2 14.49±2.45 8.08±6.19 3.64±2.22 2.95±3.33 3.57±0.87 Unigene0023040 IAA 3 213.97±40.34 119.15±61.21 132.16±54.901 48.88±36.07 39.56±9.26 Unigene0025388 IAA 4 4.61±3.19 5.45±0.97 8.84±2.12 1.18±0.84 0.00 Unigene0027417 IAA 5 11.82±1.17 7.21±3.44 5.10±1.42 1.48±1.15 3.18±0.68 Unigene0027794 IAA 6 2.28±0.33 0.82±0.67 0.53±0.42 0.28±0.39 0.00 Unigene0028701 IAA 7 60.78±12.10 30.43±21.84 77.84±20.86 50.67±17.82 21.64±5.46 Unigene0028916 IAA 8 127.67±26.48 40.53±21.48 66.59±24.93 35.91±11.76 14.06±1.25 Unigene0030426 IAA 9 7.79±1.10 2.82±3.12 3.91±1.72 0.36±0.51 2.20±0.80 与GA3合成相关基因
Genes related to GA3 synthesisUnigene0002430 GA3 1 24.60±5.23 24.72±10.21 18.71±6.22 5.74±6.31 2.92±0.83 Unigene0019924 GA3 2 4.80±1.80 0.62±0.60 1.20±1.54 0.00 0.00 Unigene0021502 GA3 3 4.02±1.56 1.21±1.06 3.05±2.99 2.34±1.78 0.00 Unigene0021503 GA3 4 5.82±3.34 5.51±2.87 5.20±2.06 6.88±7.74 0.00 Unigene0027812 GA3 5 13.12±2.80 1.38±1.23 2.59±0.96 1.91±0.92 5.69±1.14 Unigene0023907 GA3 6 16.76±10.56 4.20±5.94 0.67±0.60 0.08±0.12 0.42±0.60 Unigene0024440 GA3 7 176.89±19.66 189.53±72.98 110.84±19.67 80.80±39.49 423.10±68.62 Unigene0026445 GA3 8 47.20±12.01 32.18±34.09 51.94±10.28 74.13±97.99 203.55±60.52 Unigene0020765 GA3 9 11.95±2.51 1.74±1.95 4.61±2.35 0.55±0.78 0.00 在山药块茎的膨大过程中,GA3合成相关的基因表达量里,有2个基因的表达量为上调表达,分别是GA3 7和GA3 8,其中GA3 7的rpkm值从176.89上升至423.10,GA3 8的rpkm值从203.55上升至47.20,其余的7个基因表达量均为下调表达。
2.5 内源激素含量与其基因表达量的相关性
使用SPSS 25.0软件,对毕克齐山药的整个生长发育过程中的IAA和GA3与其合成相关基因的表达量分别进行了相关性分析(表5、6)。由表5可知,IAA含量与相关的基因IAA6呈显著负相关(P<0.05),相关系数为−0.902,结合毕克齐山药的IAA含量为上升变化(表1),而IAA6的基因表达量为下调表达(表4),相关性分析与这两个结果相符。由表6可知,GA3含量与基因GA3 1、GA3 2、GA3 3、GA3 4、GA3 5、GA3 6和GA3 9呈正相关,与GA3 7和GA3 8呈负相关,其中与GA3 5呈显著正相关(P<0.05),相关系数为0.900,结合毕克齐山药的GA3含量为先下降后上升的变化趋势(表1),而GA3 5的基因表达量也为先下调表达,后上调表达(表4),两项分析结果相符。
表 5 内源激素IAA含量与其合成相关基因表达量的相关性分析Table 5. Correlation between IAA and expression of IAA synthesis-related gene项目
ItemIAA含量
IAA contentIAA 1 IAA 2 IAA 3 IAA 4 IAA 5 IAA 6 IAA 7 IAA 8 IAA 9 IAA含量 IAA content −0.681 −0.875 −0.719 −0.046 −0.744 −0.902* −0.230 −0.799 −0.644 IAA 1 0.868 0.992** 0.669 0.956* 0.912* 0.482 0.898* 0.916* IAA 2 0.869 0.218 0.961** 0.966** 0.128 0.833 0.869 IAA 3 0.638 0.944* 0.936* 0.547 0.946* 0.936* IAA 4 0.442 0.340 0.700 0.481 0.453 IAA 5 0.944* 0.245 0.855 0.937* IAA 6 0.373 0.944* 0.904* IAA 7 0.643 0.418 IAA 8 0.911* IAA 9 表 6 内源激素GA3含量与其合成相关基因表达量的相关性分析Table 6. Correlation between GA3 and expression of GA3 synthesis-related gene项目
ItemGA3含量
GA3 contentGA3 1 GA3 2 GA3 3 GA3 4 GA3 5 GA3 6 GA3 7 GA3 8 GA3 9 GA3含量 GA3 content 0.256 0.863 0.680 0.020 0.900* 0.737 −0.007 −0.041 0.872 GA3 1 0.652 0.557 0.511 0.267 0.648 −0.400 −0.803 0.673 GA3 2 0.765 0.306 0.876 0.958* −0.185 −0.416 0.991** GA3 3 0.728 0.468 0.620 −0.730 −0.704 0.826 GA3 4 −0.096 0.277 −0.957* −0.910* 0.354 GA3 5 0.873 0.224 0.058 0.827 GA3 6 −0.085 −0.383 0.914* GA3 7 0.854 −0.265 GA3 8 −0.469 GA3 9 3. 讨论
3.1 块茎生理指标的变化
毕克齐山药的可溶性总糖含量在块茎膨大前期缓慢下降,块茎膨大中期和后期基本保持不变;还原糖含量在块茎膨大前期和中期均匀下降,块茎膨大后期略有上升;淀粉含量在块茎膨大前期快速上升,此时糖转化为淀粉慢慢积累,之后为生长提供能量所以快速下降至第四时间段,块茎膨大后期时淀粉含量缓慢上升,由于此时山药块茎发育变得较为缓慢,所以一部分糖除了供能外其余转化为淀粉进行储存。
块茎可溶性总糖和还原糖含量表现为下降趋势,淀粉含量均表现为上升趋势,并且淀粉含量与还原糖含量和可溶性总糖含量表现出负相关关系,说明在山药整个生长发育过程中,还原糖和可溶性总糖一部分为山药生长提供能量被消耗,另一部分转化为淀粉,淀粉在整个过程中持续积累,说明了淀粉与糖之间的互相转化。孙霞[14]在对毕克齐山药生长发育期间的营养成分研究发现,可溶性总糖含量和还原糖含量与淀粉含量呈极显著负相关。本研究结果与孙霞研究结果一致。
3.2 内源激素含量对块茎膨大的影响
在毕克齐山药块茎膨大过程中,IAA含量在种植后105~135 d 时上升,之后下降至种植后150 d,再上升至种植后165 d,结合其形态特点为“细长”型山药,原因可能是为促进山药的伸长生长,IAA含量第二次出现上升,或者是因为内源激素本身较为敏感,在取样或试验过程中造成的试验误差所致。IAA含量在块茎开始形成时含量逐渐上升,直至135 d达到最高,之后开始下降,这与李明军[15]研究结果一致。
ZR作为细胞分裂素的主要组成成分,主要通过促进细胞分裂、抑制细胞伸长、促进细胞扩展来影响甘薯块根形成和膨大[16]。本研究中,ZR含量在毕克齐山药的整个膨大期中表现出先升后降趋势,块茎膨大前期细胞分裂,块茎内产生大量ZR来促进细胞分裂,块茎膨大后期,块茎发育成熟,细胞分裂减慢,ZR含量下降来调整细胞分裂速度,这与李良俊[17]等人研究结果一致。
ABA含量在块茎膨大前期至块茎膨大中期无明显变化,块茎膨大后期时ABA含量快速升高并达到最大值,因为ABA主要合成于处在休眠状态和将要脱落的器官中[18],所以在山药块茎膨大后期ABA含量快速升高。本研究结果与李良俊[17]、龙雯虹[19]等的研究结果一致。
茉莉酸(JA)和茉莉酸甲酯(MeJA)及其衍生物对马铃薯块茎发育和诱导有重要作用,对植物根的生长,果实成熟等生长发育过程有重要调控作用[20]。JA促进马铃薯鲜重、直径的增加,并且低浓度的JA含量促进马铃薯块茎体积增大[21]。本研究中,毕克齐山药的JA含量在整个块茎膨大过程中波动上升,在块茎膨大后期表现为上升趋势,此时期JA促进山药衰老、块茎成熟。
马铃薯块茎形成的过程中受到多种调控途径的调控,其中赤霉素(GA)途径是最重要的途径之一[22]。本研究中,毕克齐山药的GA3含量在块茎膨大前期下降,说明GA3含量降低块茎开始形成,本研究中GA3在种植后120~135 d上升,种植后135~165 d下降,此时山药块茎发育逐渐成熟。龚明霞[23]等研究发现,GA3浓度过高会产生负效应,不利于增产。本研究结果与其结果一致。
本研究中,IPA含量在块茎膨大前期快速上升,此时期的细胞分裂旺盛,块茎开始膨大,IPA作为细胞分裂素的一种,含量上升,促进细胞分裂,而IPA含量在块茎膨大中期下降至块茎膨大后期,此时块茎生长节奏放缓,细胞分裂不旺盛,IPA含量下降。
水杨酸(SA)参与调节植物的种子萌发、开花等生长发育相关的生理过程,近年来研究人员也发现水杨酸在提高植物抗热、抗寒、抗盐、抗旱等方面具有重要作用[24]。本研究中,毕克齐山药的SA含量在块茎膨大后期均表现为快速上升趋势,可能是由于这段时期内蒙古地区进入深秋季节,气温下降迅速且昼夜温差大,同时地面温度和地下温度也在下降,山药通过自身调节,使SA含量上升从而应对较低的温度,本研究结果与李良俊[17]研究结果类似。
毕克齐山药在整个块茎膨大过程中,内源激素的变化存在差异,这可能是由于这7种内源激素对山药调控的时序性不同,也可能是由于山药本身的生理活动以及各种代谢的差异所引起。
3.3 内源激素含量与生理指标的相关性
毕克齐山药中7种内源激素与各生理指标的相关性分析表明,内源激素IAA、ZR、ABA和JA含量与淀粉含量呈正相关,可能因为生长素和细胞分裂素影响光合产物的积累和运输,从而促进块茎中淀粉含量的积累,GA3、IPA和SA与淀粉含量呈负相关,这些激素可能影响了山药的成熟和衰老,杨鑫[25]等人对木薯块根中内源激素含量与淀粉积累关系的研究发现,木薯块根中GA3与淀粉含量呈极显著正相关关系,本研究中GA3与形态指标呈负相关,其中与块茎长度呈显著负相关,本研究结果与杨鑫研究结果不一致;除GA3与还原糖含量呈正相关之外,其余6种内源激素均与还原糖含量呈负相关,其中ZR与其呈显著负相关,可能是玉米素通过影响发育过程中的转化酶活性来影响糖分积累[26];IAA、ZR、ABA、JA、IPA和SA分别与可溶性总糖含量呈负相关,其中IAA与其呈显著负相关,苏艳[27]在对草莓果实糖代谢规律的研究中发现,花托中的蔗糖和葡萄糖及瘦果中的葡萄糖和果糖对生长素信号响应较强,本研究中IAA与可溶性总糖含量呈显著负相关,所以本研究结果与苏艳研究结果均表明生长素与糖含量的较强关联性。
3.4 内源激素与其相关基因表达量的关系
对毕克齐山药转录组数据分析与筛选,获得了与IAA相关的基因和与GA3相关的基因。
IAA含量的变化随着毕克齐山药块茎的生长发育不断升高,在整个块茎膨大过程中其含量呈上升趋势,而与IAA相关的基因表达量均为下调表达,说明IAA相关基因正调控IAA含量。
生长素参与植物的生长发育,并且发挥着重要作用。生长素响应蛋白在生长素信号转导过程起着十分关键的作用[28]。罗莎[29]对拟南芥的研究表明,生长素响应蛋白AUX/IAA能调节植物发育以及生长素诱导基因的表达,并且生长素响应需要AUX/IAA蛋白周转。王益军等[30]在高粱全基因组的研究中发现:AUX/IAA基因家族对生长素早期诱导具有响应作用,并且AUX/IAA蛋白作为一类逆转录因子[31],对生长素具有负调控作用。本研究中,毕克齐山药在整个生长发育过程中IAA含量呈上升趋势,与IAA相关的9个基因中7个生长素响应蛋白基因:IAA1、IAA2、IAA3、IAA6、IAA7、IAA8和IAA9的表达量均为下调表达,相关性分析结果也表明,IAA含量和IAA6基因表达量呈显著负相关,在毕克齐山药中,这7个生长素响应蛋白基因的下调表达,对IAA有正向调控作用。本研究结果与以上研究结果一致。
Hongqi Du等[32]在对苜蓿秋季休眠的研究中发现,IAA-氨基酸水解酶(IAA-amino acid hydrolase ILR1-like1)参与调节苜蓿的秋季休眠。本研究在毕克齐山药块茎中发现了与IAA相关的基因,其中有2个IAA-氨基酸水解酶ILR1基因:IAA4和IAA5,说明IAA-氨基酸水解酶ILR1参与了IAA的合成。本研究中,IAA4的表达量在第105 d至第135 d上升,可能是该基因参与促进毕克齐山药中IAA含量的升高。
本研究中,GA3含量在毕克齐山药块茎的整个膨大过程中表现为波动变化,整体呈下降趋势,而与GA3合成相关的基因,除了GA37和GA38的表达量为上升变化外,其余基因的表达量均为下降。
赤霉素是一种能够调节植物生长发育各个环节的植物激素,它可以调节包括种子萌发、下胚轴和茎的伸长以及花发育等许多方面,作用于高等植物的整个生命周期。程红亮[33]在玉米叶色突变基因定位中发现,赤霉素2-β-双加氧酶是赤霉素灭活中的关键酶。本研究中,毕克齐山药GA3含量(表1)在块茎膨大前期之后迅速下降,与GA3相关的赤霉素2-β-双加氧酶:GA3 1、GA3 2、GA3 3和GA3 4,这些基因在块茎膨大前期也呈下降趋势,可能赤霉素2-β-双加氧酶参与调控了赤霉素灭活,使其含量下降。
赤霉素20氧化酶(Gibberellin 20 oxidase, GA20ox)是合成赤霉素的关键限速酶,GA20ox通过调节赤霉素的合成及其含量,来调控植物生长发育的各个阶段[34]。本研究中,毕克齐山药的GA3含量变化趋势为先下降后上升再下降,这与GA3 5的表达量变化一致,同时结合相关性分析,GA3含量与GA3 5呈显著正相关,可能说明赤霉素的合成与赤霉素20氧化酶密不可分,本研究结果与上述研究结果类似。
孙雷明[35]在柑橘成花发育分子机制的研究中发现,赤霉素调节蛋白参与了激素调控,所以推测在本研究中,毕克齐山药中的GA3受赤霉素调节蛋白基因GA3 6、GA3 7、GA3 8和GA3 9调控。
4. 结论
山药块茎内部的淀粉、还原糖和可溶性总糖在合成、运转及积累过程中均存在一定的相关性,在整个生长发育过程中,块茎淀粉含量持续增加,还原糖和可溶性总糖含量在整个块茎膨大期间逐渐下降。
内源激素ZR、ABA、SA与山药块茎长度、块茎周长、块茎直径、块茎鲜重、块茎干重正相关,即促进山药块茎膨大;IPA与各形态指标负相关,抑制山药块茎生长;IAA与山药块茎周长和块茎直径显著正相关,促进山药增粗;GA3与块茎长度显著负相关,抑制其伸长生长。
通过分析与内源激素相关的差异表达基因,结果表明:IAA和与合成其相关的基因IAA 6显著负相关,相关基因IAA 6对IAA 起负调控作用;GA3和与合成其相关的基因GA3 5显著正相关,相关基因GA3 5正向调控GA3。
-
![]()
图 1 生长发育过程各个生理指标的变化
注:A-生长发育过程还原糖含量的变化;B-生长发育过程可溶性总糖含量的变化;C-生长发育过程淀粉含量的变化。
Figure 1. Physiological changes on Bikeqi yam during tuber development
Note: A-on reducing sugars; B-on soluble sugars; C-on starch.
表 1 毕克齐山药在块茎膨大过程内源激素含量变化
Table 1 Changes on endogenous hormones during tuber enlargement of Bikeqi yam
[单位: ng·g−1(FW)] 项目 Item 105 d 120 d 135 d 150 d 165 d 生长素 IAA 24.697±1.82 b 38.542±1.76 b 46.454±3.46 a 37.793±2.41 b 48.012±2.68 a 玉米素 ZR 5.295±0.23 b 6.97±0.39b 8.129±0.45 ab 9.39±0.72 a 6.751±0.36 b 脱落酸 ABA 33.274±1.93 bc 33.564±0.03 bc 28.402±1.70 c 40.707±2.11 b 81.939±1.88 a 茉莉酸 JA 11.976±0.43 b 12.678±0.39 b 15.806±1.02 ab 13.541±0.92 b 21.276±1.46 a 赤霉素 GA3 5.484±0.44 a 2.538±0.09 c 4.072±0.36 bc 3.209±0.20 bc 3.69±0.26 bc 异戊烯基腺苷 IPA 4.103±0.20 b 8.005±0.30 a 5.943±0.37 ab 4.202±0.21 b 4.419±0.27 b 水杨酸 SA 52.308±30.38 b 88.763±63.26 ab 19.226±10.83 b 81±37.48 ab 234.867±111.44 a 注:同行不同小写字母表示在0.05水平上差异显著。
Note: Data with different lowercase letters indicate significant differences at 0.05 level. 下载: 导出CSV
下载: 导出CSV
表 2 毕克齐山药内源激素与形态、生理指标相关性
Table 2 Correlation between endogenous hormones and morphology and physiology of Bikeqi yam tubers
项目
Item块茎长度
Length块茎周长
Circumference块茎直径
Diameter块茎鲜重
Fresh
weight块茎干重
Dry
weight淀粉含量
Starch
content还原糖含量
Reducing sugar
content可溶性总糖含量
Total soluble
sugar contentIAA ZR ABA JA GA3 IPA SA 块茎长度
Length0.756 0.757 0.674 0.679 0.375 −0.78 −0.940* 0.841 0.562 0.431 0.531 −0.892* 0.435 0.494 块茎周长
Circumference1.000** 0.922* 0.981** 0.719 −0.823 −0.839 0.906* 0.589 0.64 0.833 −0.425 −0.161 0.553 块茎直径
Diameter0.924* 0.982** 0.715 −0.822 −0.838 0.904* 0.588 0.643 0.833 −0.425 −0.162 0.557 块茎鲜重
Fresh weight0.972** 0.489 −0.84 −0.75 0.692 0.657 0.665 0.694 −0.43 −0.362 0.591 块茎干重
Dry weight0.646 −0.817 −0.771 0.807 0.602 0.677 0.804 −0.36 −0.321 0.579 淀粉含量
Starch content−0.617 −0.611 0.767 0.542 0.072 0.524 −0.061 −0.04 −0.082 还原糖含量
Reducing sugar content0.927* −0.732 −0.932* −0.21 −0.375 0.673 −0.034 −0.170 可溶性总糖含量
Total soluble sugar content−0.883* −0.773 −0.289 −0.494 0.805 −0.315 −0.293 IAA 0.483 0.479 0.779 −0.533 0.227 0.433 ZR −0.116 0.043 −0.575 0.022 −0.159 ABA 0.871 −0.118 −0.378 0.975** JA −0.107 −0.242 0.787 GA3 −0.612 −0.252 IPA −0.222 注:*在0.05水平,相关性显著。**在0.01水平,相关性极显著。表5、6同。
Note:*Significant correlation (P<0.05); **Extremely significant correlation (P<0.01). Same for Tables 5 and 6.
下载: 导出CSV
表 3 与内源激素合成相关基因的基因编号、基因代号和基因注释信息
Table 3 Number, code, and information on genes associated with endogenous hormone synthesis in Bikeqi yam tubers
项目
Item基因
ID Gene ID基因代号
Gene code基因注释信息
Gene annotation information与IAA合成相关基因 Genes related to IAA synthesis Unigene0016848 IAA 1 生长素响应蛋白基因 Auxin response protein gene Unigene0021936 IAA 2 生长素响应蛋白基因 Auxin response protein gene Unigene0023040 IAA 3 生长素响应蛋白基因 Auxin response protein gene Unigene0025388 IAA 4 IAA-氨基酸水解酶 ILR1基因 IAA-amino acid hydrolase ILR1 gene Unigene0027417 IAA 5 IAA-氨基酸水解酶 ILR1基因 IAA-amino acid hydrolase ILR1 gene Unigene0027794 IAA 6 生长素响应蛋白基因 Auxin response protein gene Unigene0028701 IAA 7 生长素响应蛋白基因 Auxin response protein gene Unigene0028916 IAA 8 生长素响应蛋白基因 Auxin response protein gene Unigene0030426 IAA 9 生长素响应蛋白基因 Auxin response protein gene 与 GA3合成相关基因 Genes related to GA3 synthesis Unigene0002430 GA3 1 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0019924 GA3 2 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0021502 GA3 3 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0021503 GA3 4 赤霉素2-β-双加氧酶 Gibberellin 2-β-dioxygenase Unigene0027812 GA3 5 赤霉素20氧化酶 Gibberellin 2-β-dioxygenase Unigene0023907 GA3 6 赤霉素调节蛋白基因 Gibberellin regulatory protein gene Unigene0024440 GA3 7 赤霉素调节蛋白基因 Gibberellin regulatory protein gene Unigene0026445 GA3 8 赤霉素调节蛋白基因 Gibberellin regulatory protein gene Unigene0020765 GA3 9 赤霉素调节蛋白基因 Gibberellin regulatory protein gene
下载: 导出CSV
表 4 与内源激素合成相关基因的rpkm平均值
Table 4 Average rpkm of genes associated with endogenous hormone synthesis
项目
Item基因ID
Gene ID基因代号
Gene code105 d 120 d 135 d 150 d 165 d 与IAA合成相关基因
Genes related to IAA synthesisUnigene0016848 IAA 1 95.82±30.63 65.29±44.90 64.25±22.21 26.73±12.83 26.68±10.96 Unigene0021936 IAA 2 14.49±2.45 8.08±6.19 3.64±2.22 2.95±3.33 3.57±0.87 Unigene0023040 IAA 3 213.97±40.34 119.15±61.21 132.16±54.901 48.88±36.07 39.56±9.26 Unigene0025388 IAA 4 4.61±3.19 5.45±0.97 8.84±2.12 1.18±0.84 0.00 Unigene0027417 IAA 5 11.82±1.17 7.21±3.44 5.10±1.42 1.48±1.15 3.18±0.68 Unigene0027794 IAA 6 2.28±0.33 0.82±0.67 0.53±0.42 0.28±0.39 0.00 Unigene0028701 IAA 7 60.78±12.10 30.43±21.84 77.84±20.86 50.67±17.82 21.64±5.46 Unigene0028916 IAA 8 127.67±26.48 40.53±21.48 66.59±24.93 35.91±11.76 14.06±1.25 Unigene0030426 IAA 9 7.79±1.10 2.82±3.12 3.91±1.72 0.36±0.51 2.20±0.80 与GA3合成相关基因
Genes related to GA3 synthesisUnigene0002430 GA3 1 24.60±5.23 24.72±10.21 18.71±6.22 5.74±6.31 2.92±0.83 Unigene0019924 GA3 2 4.80±1.80 0.62±0.60 1.20±1.54 0.00 0.00 Unigene0021502 GA3 3 4.02±1.56 1.21±1.06 3.05±2.99 2.34±1.78 0.00 Unigene0021503 GA3 4 5.82±3.34 5.51±2.87 5.20±2.06 6.88±7.74 0.00 Unigene0027812 GA3 5 13.12±2.80 1.38±1.23 2.59±0.96 1.91±0.92 5.69±1.14 Unigene0023907 GA3 6 16.76±10.56 4.20±5.94 0.67±0.60 0.08±0.12 0.42±0.60 Unigene0024440 GA3 7 176.89±19.66 189.53±72.98 110.84±19.67 80.80±39.49 423.10±68.62 Unigene0026445 GA3 8 47.20±12.01 32.18±34.09 51.94±10.28 74.13±97.99 203.55±60.52 Unigene0020765 GA3 9 11.95±2.51 1.74±1.95 4.61±2.35 0.55±0.78 0.00
下载: 导出CSV
表 5 内源激素IAA含量与其合成相关基因表达量的相关性分析
Table 5 Correlation between IAA and expression of IAA synthesis-related gene
项目
ItemIAA含量
IAA contentIAA 1 IAA 2 IAA 3 IAA 4 IAA 5 IAA 6 IAA 7 IAA 8 IAA 9 IAA含量 IAA content −0.681 −0.875 −0.719 −0.046 −0.744 −0.902* −0.230 −0.799 −0.644 IAA 1 0.868 0.992** 0.669 0.956* 0.912* 0.482 0.898* 0.916* IAA 2 0.869 0.218 0.961** 0.966** 0.128 0.833 0.869 IAA 3 0.638 0.944* 0.936* 0.547 0.946* 0.936* IAA 4 0.442 0.340 0.700 0.481 0.453 IAA 5 0.944* 0.245 0.855 0.937* IAA 6 0.373 0.944* 0.904* IAA 7 0.643 0.418 IAA 8 0.911* IAA 9
下载: 导出CSV
表 6 内源激素GA3含量与其合成相关基因表达量的相关性分析
Table 6 Correlation between GA3 and expression of GA3 synthesis-related gene
项目
ItemGA3含量
GA3 contentGA3 1 GA3 2 GA3 3 GA3 4 GA3 5 GA3 6 GA3 7 GA3 8 GA3 9 GA3含量 GA3 content 0.256 0.863 0.680 0.020 0.900* 0.737 −0.007 −0.041 0.872 GA3 1 0.652 0.557 0.511 0.267 0.648 −0.400 −0.803 0.673 GA3 2 0.765 0.306 0.876 0.958* −0.185 −0.416 0.991** GA3 3 0.728 0.468 0.620 −0.730 −0.704 0.826 GA3 4 −0.096 0.277 −0.957* −0.910* 0.354 GA3 5 0.873 0.224 0.058 0.827 GA3 6 −0.085 −0.383 0.914* GA3 7 0.854 −0.265 GA3 8 −0.469 GA3 9
下载: 导出CSV
-
[1] 李艳英, 甘秀芹, 韦本辉, 等. 64份淮山种质资源品质性状分析 [J]. 植物遗传资源学报, 2016, 17(2):246−251. LI Y Y, GAN X Q, WEI B H, et al. Analysis on quality characters of 64 yam(Dioscorea L.) germplasm resources [J]. Journal of Plant Genetic Resources, 2016, 17(2): 246−251.(in Chinese
[2] SANTNER A, CALDERON-VILLALOBOS L I A, ESTELLE M. Plant hormones are versatile chemical regulators of plant growth [J]. Nature Chemical Biology, 2009, 5(5): 301−307. DOI: 10.1038/nchembio.165
[3] 李玲玲. 菊芋块茎形成及其与内源激素的关系初步研究[D]. 南京: 南京农业大学, 2015. LI L L. Preliminary researches on Tuber formation of Helianthus tubersosus land the relation with endogenous phytohormones[D]. Nanjing: Nanjing Agricultural University, 2015. (in Chinese).
[4] 王志敏, 霍秀文, 张艳芳, 等. 山药(Dioscorea opposita)块茎不同发育时期相关差异基因的cDNA-AFLP分析 [J]. 分子植物育种, 2019, 17(11):3650−3659. WANG Z M, HUO X W, ZHANG Y F, et al. cDNA-AFLP analysis of differential genes related to different development stages of Dioscorea opposita tubers [J]. Molecular Plant Breeding, 2019, 17(11): 3650−3659.(in Chinese
[5] 梁任繁, 李创珍, 张娟, 等. 山药块茎发育中物质积累及相关代谢酶变化 [J]. 作物学报, 2011, 37(5):903−910. DOI: 10.3724/SP.J.1006.2011.00903 LIANG R F, LI C Z, ZHANG J, et al. Changes of matter accumulation and relative enzymatic activity during yam Tuber development [J]. Acta Agronomica Sinica, 2011, 37(5): 903−910.(in Chinese DOI: 10.3724/SP.J.1006.2011.00903
[6] 周芸伊. 山药DELLA蛋白参与赤霉素调控块茎生长膨大的分子机制[C]//2017年中国作物学会学术年会论文摘要集, 2017, 130-131. [7] 周芸伊, 张静, 王亚伦, 等. 赤霉素调控植物块茎形态建成的研究进展 [J]. 作物杂志, 2016(4):20−25. ZHOU Y Y, ZHANG J, WANG Y L, et al. Progress of gibberellin regulation on Tuber morphogenesis in higher plant [J]. Crops, 2016(4): 20−25.(in Chinese
[8] 甘立军, 曾晓春, 周燮. 茉莉酸类与植物地下贮藏器官的形成 [J]. 植物学通报, 2001, 18(5):546−553. GAN L J, ZENG X C, ZHOU X. Possible involvement of jasmonates in the morphogenesis of underground storage organs in plants [J]. Chinese Bulletin of Botany, 2001, 18(5): 546−553.(in Chinese
[9] 汪雷, 马琛, 高海立, 等. 钙对半夏生理特性及光合生理的影响 [J]. 浙江理工大学学报(自然科学版), 2018, 39(4):461−467. WANG L, MA C, GAO H L, et al. Effects of calcium on physiological characteristics and photosynthetic physiology of Pinellia ternata [J]. Journal of Zhejiang Institute of Science and Technology, 2018, 39(4): 461−467.(in Chinese
[10] ROUMELIOTIS E, KLOOSTERMAN B, OORTWIJN M, et al. The effects of auxin and strigolactones on Tuber initiation and stolon architecture in potato [J]. Journal of Experimental Botany, 2012, 63(12): 4539−4547. DOI: 10.1093/jxb/ers132
[11] ZHANG Z J, ZHOU W J, LI H Z. The role of GA, IAA and BAP in the regulation of in vitro shoot growth and microtuberization in potato [J]. Acta Physiologiae Plantarum, 2005, 27(3): 363−369. DOI: 10.1007/s11738-005-0013-7
[12] 敖兰吉亚, 季祥, 邵盈, 等. 山药块茎生长期5种内源激素含量变化对块茎膨大的影响 [J]. 福建农业学报, 2019, 34(3):284−292. AOLAN J Y, JI X, SHAO Y, et al. Correlation between endogenous hormones and
Tuber growth of Dioscorea opposite thunb [J]. Fujian Journal of Agricultural Sciences, 2019, 34(3): 284−292.(in Chinese [13] PAN X Q, WELTI R, WANG X M. Simultaneous quantification of major phytohormones and related compounds in crude plant extracts by liquid chromatography-electrospray tandem mass spectrometry [J]. Phytochemistry, 2008, 69(8): 1773−1781. DOI: 10.1016/j.phytochem.2008.02.008
[14] 孙霞. 毕克齐长山药生育和贮藏期间营养成分及相关酶活性的研究[D]. 呼和浩特: 内蒙古农业大学, 2008. SUN X. Study on the Tuber nutrition content and pertinent enzyme activity of bikeqi yam during growth development and storage period[D]. Hohhot: Inner Mongolia Agricultural University, 2008. (in Chinese).
[15] 李明军, 刘世宇, 刘雯, 等. 怀山药微型块茎形成过程中的生理生化变化 [J]. 植物生理学报, 2017, 53(5):807−814. LI M J, LIU S Y, LIU W, et al. Physiological and biochemical changes in Dioscorea opposita during the process of microtuber formation [J]. Plant Physiology Communications, 2017, 53(5): 807−814.(in Chinese
[16] 陈小琴. 人参榕块根膨大机理及其调控技术研究[D]. 福州: 福建农林大学, 2009. CHEN X Q. Study on the enlargement mechanism and regulation technology of tuberous root of Ficus microcarpa l. f.[D]. Fuzhou: Fujian Agriculture and Forestry University, 2009. (in Chinese).
[17] 李良俊, 潘恩超, 许超, 等. 莲藕膨大过程中内源激素、水杨酸和多胺含量的变化 [J]. 园艺学报, 2006, 33(5):1106−1108. LI L J, PAN E C, XU C, et al. Changes of endogenous hormones, polyamines and salicylic acid content during rhizome development of Nelumbo nucifera gaertn [J]. Acta Horticulturae Sinica, 2006, 33(5): 1106−1108.(in Chinese
[18] 孙鹏. 植物激素对甜菜块根增长和糖分积累的调控作用[D]. 呼和浩特: 内蒙古农业大学, 2014. SUN P. The regulation of plant hormone on the increase of Tuber weight and sugar accumulation of sugar beet[D]. Hohhot: Inner Mongolia Agricultural University, 2014. (in Chinese).
[19] 龙雯虹, 郭华春, 肖关丽, 等. 山药珠芽生长过程中激素和糖类物质含量的变化 [J]. 园艺学报, 2011, 38(4):753−760. LONG W H, GUO H C, XIAO G L, et al. Variation of endogenous hormone and carbohydrate contents in growing yam bulbils [J]. Acta Horticulturae Sinica, 2011, 38(4): 753−760.(in Chinese
[20] 张培安, 左倩倩, 董天宇, 等. 茉莉酸甲酯对葡萄植株不定根发育的影响 [J]. 园艺学报, 2018, 45(12):2331−2346. ZHANG P A, ZUO Q Q, DONG T Y, et al. Effects of MeJA on adventitious root development of grape plants [J]. Acta Horticulturae Sinica, 2018, 45(12): 2331−2346.(in Chinese
[21] 王润润. 马铃薯块茎离体发育过程茉莉酸调控的差异蛋白质组分析[D]. 兰州: 甘肃农业大学, 2017. WANG R R. Comparative proteomic analysis of potato Tuber development in vitro regulated by jasmonate acid[D]. Lanzhou: Gansu Agricultural University, 2017. (in Chinese).
[22] 何依雪, 刘文, 沈祥陵. 蔗糖, GA与液体培养基对马铃薯块茎形成的影响 [J]. 生物技术通报, 2018, 34(7):74−80. HE Y X, LIU W, SHEN X L. Effect of sucrose, GA and liquid medium on the Tuber formation of potato [J]. Biotechnology Bulletin, 2018, 34(7): 74−80.(in Chinese
[23] 龚明霞, 罗海玲, 袁红娟, 等. 外源赤霉素和多效唑对山药块茎膨大和零余子形成的影响 [J]. 园艺学报, 2015, 42(6):1175−1184. GONG M X, LUO H L, YUAN H J, et al. Effects of exogenous gibberellin and paclobutrazol on Tuber expansion and bulbil formation of Chinese yam [J]. Acta Horticulturae Sinica, 2015, 42(6): 1175−1184.(in Chinese
[24] 李亮. 水杨酸在黄瓜(Cucumis sativus L.)幼苗应答低温胁迫中的作用机制[D]. 北京: 中国农业科学院, 2013. LI L. Roles of salicylic acid in response to low temperature stress in cucumber (Cucumis sativus L.) seedlings[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. (in Chinese).
[25] 杨鑫, 罗兴录, 覃宏宇, 等. 木薯内源激素含量与块根淀粉积累关系研究 [J]. 中国农学通报, 2013, 29(33):158−164. YANG X, LUO X L, QIN H Y, et al. Study on endogenous hormones content and its relations to tuberous root starch accumulation in cassava [J]. Chinese Agricultural Science Bulletin, 2013, 29(33): 158−164.(in Chinese
[26] 肖年湘, 郁松林, 王春飞. 6-BA、玉米素对全球红葡萄果实发育过程中糖分含量和转化酶活性的影响 [J]. 西北农业学报, 2008, 17(3):227−231. XIAO N X, YU S L, WANG C F. Effects of 6-BA and Zeatin on the fruit sugar contents and invertase activities in red globe grape during fruit development [J]. Acta Agriculturae Boreali-Occidentalis Sinica, 2008, 17(3): 227−231.(in Chinese
[27] 苏艳. 草莓果实糖代谢规律及其对生长素信号的响应[D]. 北京: 北京林业大学, 2009. SU Y. Study on the regulation of sugar metabolism and sugar metabolism respond to auxin signaling in developing fruits of strawberry[D]. Beijing: Beijing Forestry University, 2009. (in Chinese).
[28] 张丽丽. 三个棉花生长素信号转导途径相关基因的克隆与鉴定[D]. 南京: 南京农业大学, 2010. ZHANG L L. Cloning and characterization of three genes related with auxin signal transduction in Gossypium hirsutum l.[D]. Nanjing: Nanjing Agricultural University, 2010. (in Chinese).
[29] 罗莎. Aux/IAA家族转录因子OsIAA9对拟南芥生长素信号转导及根部生长发育的调控[D]. 长春: 东北师范大学, 2015. LUO S. Regulation of auxin signaling and root growth by Aux/IAA transcription factor OsIAA9 in Arabidopsis[D]. Changchun: Northeast Normal University, 2015. (in Chinese).
[30] 王益军, 吕燕萍, 谢秦, 等. 高粱全基因组生长素原初响应基因Aux/IAA的序列特征分析 [J]. 作物学报, 2010, 36(4):688−694. DOI: 10.3724/SP.J.1006.2010.00688 WANG Y J, LV Y P, XIE Q, et al. Whole-genome sequence characterization of primary auxin-responsive Aux/IAA gene family in Sorghum (Sorghum bicolor L.) [J]. Acta Agronomica Sinica, 2010, 36(4): 688−694.(in Chinese DOI: 10.3724/SP.J.1006.2010.00688
[31] 林伟强. 拟南芥Aux/IAA家族基因IAA2的反向遗传学功能研究[D]. 杭州: 浙江大学, 2007. LIN W Q. Functional analyses of Aux/IAA family member IAA2 by reverse genetic approaches in Arabidopsis thaliana[D]. Hangzhou: Zhejiang University, 2007. (in Chinese).
[32] DU H Q, SHI Y H, LI D F, et al. Screening and identification of key genes regulating fall dormancy in alfalfa leaves [J]. US National Library of Medicine National Institutes of Health, 2017, 12(2): 1−25.
[33] 程红亮, 陈甲法, 丁俊强, 等. 一个玉米叶色突变体的遗传分析与基因定位 [J]. 华北农学报, 2011, 26(3):7−10. CHENG H L, CHEN J F, DING J Q, et al. Genetic analysis and gene mapping of a leaf mutant in maize [J]. Acta Agriculturae Boreali-Sinica, 2011, 26(3): 7−10.(in Chinese
[34] 吴建明, 陈荣发, 黄杏, 等. 高等植物赤霉素生物合成关键组分 GA20-oxidase 氧化酶基因的研究进展 [J]. 生物技术通报, 2016, 32(7):1−12. WU J M, CHEN R F, HUANG X, et al. Studies on the gene of key component GA20-oxidase for gibberellin biosynthesis in plant [J]. Biotechnology Bulletin, 2016, 32(7): 1−12.(in Chinese
[35] SUN L M, AI X Y, LI W Y, et al. Identification and comparative profiling of miRNAs in an early flowering mutant of trifoliate orange and its wild type by genome-wide deep sequencing [J]. PLoS One, 2012, 7(8): e43760. DOI: 10.1371/journal.pone.0043760
-
期刊类型引用(4)
1. 王慧娟,王二强,符真珠,李艳敏,王晓晖,袁欣,高杰,王利民,张和臣. 芍药根茎形成发育过程中内源激素和碳水化合物的变化. 河南农业科学. 2024(03): 118-124 .  百度学术
百度学术
2. 刘德才,金林,郭文琦,殷剑美,王立,蒋璐,韩晓勇,张培通. 渍水对定向槽种植薯蓣类山药块茎发育的影响. 安徽农业科学. 2024(18): 158-161 . 百度学术
3. 梁任繁,黄皓,周生茂,王爱勤,詹洁,文俊丽,何龙飞. 山药块茎生长膨大相关基因的筛选与分析. 中国蔬菜. 2024(09): 61-67 . 百度学术
4. 周春艳,狄永国,仇全雷,蔡金龙,刘云霞,徐慧妮,李昆志. 转录组和代谢组联合分析低温胁迫对天麻生长发育的影响. 分子植物育种. 2023(01): 110-122 . 百度学术
其他类型引用(4)
计量
- 文章访问数: 971
- HTML全文浏览量: 262
- PDF下载量: 21
- 被引次数: 8
