

Codon Usage Bias of Chloroplast Genome in Liparis bootanensis
-
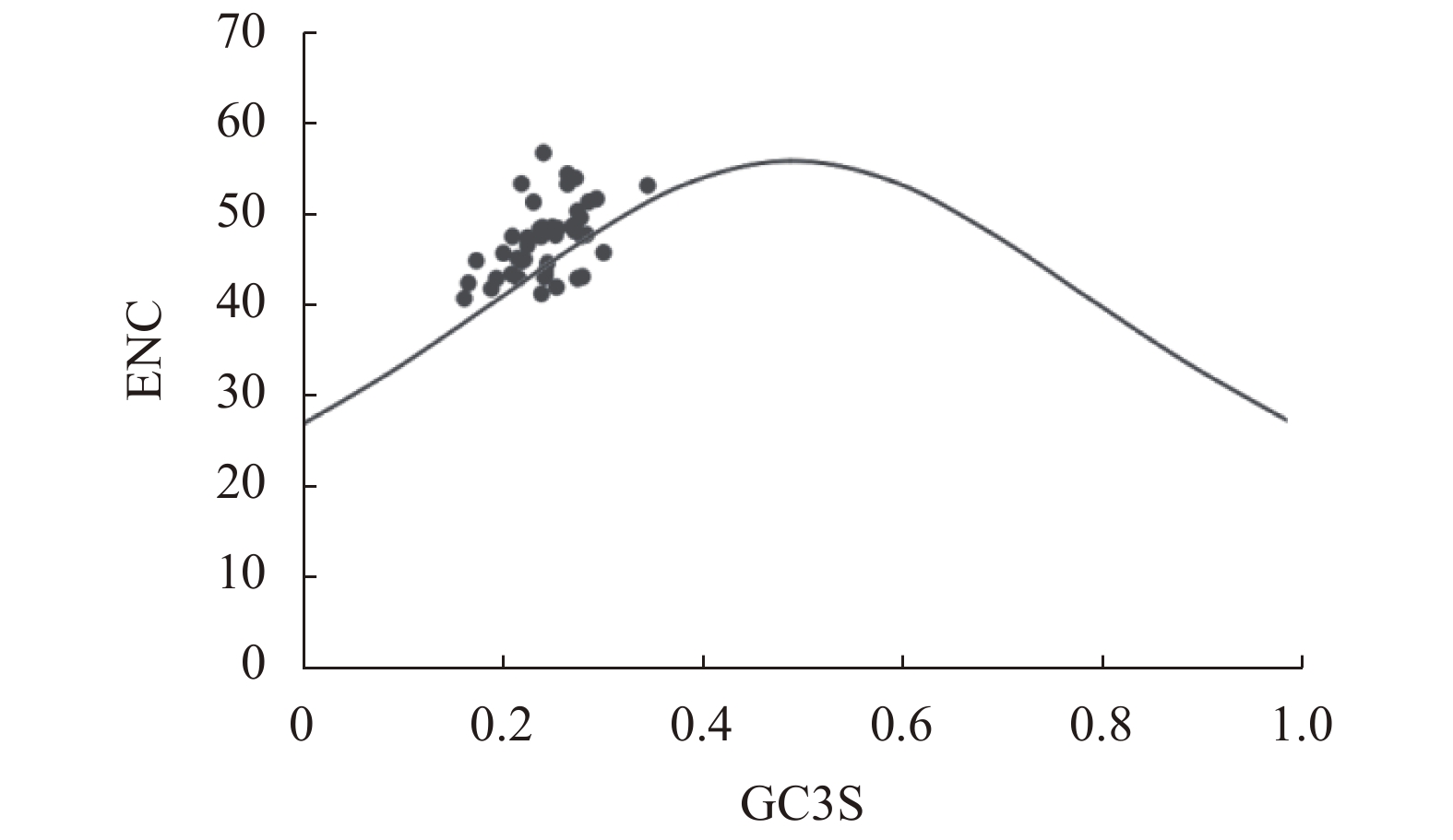
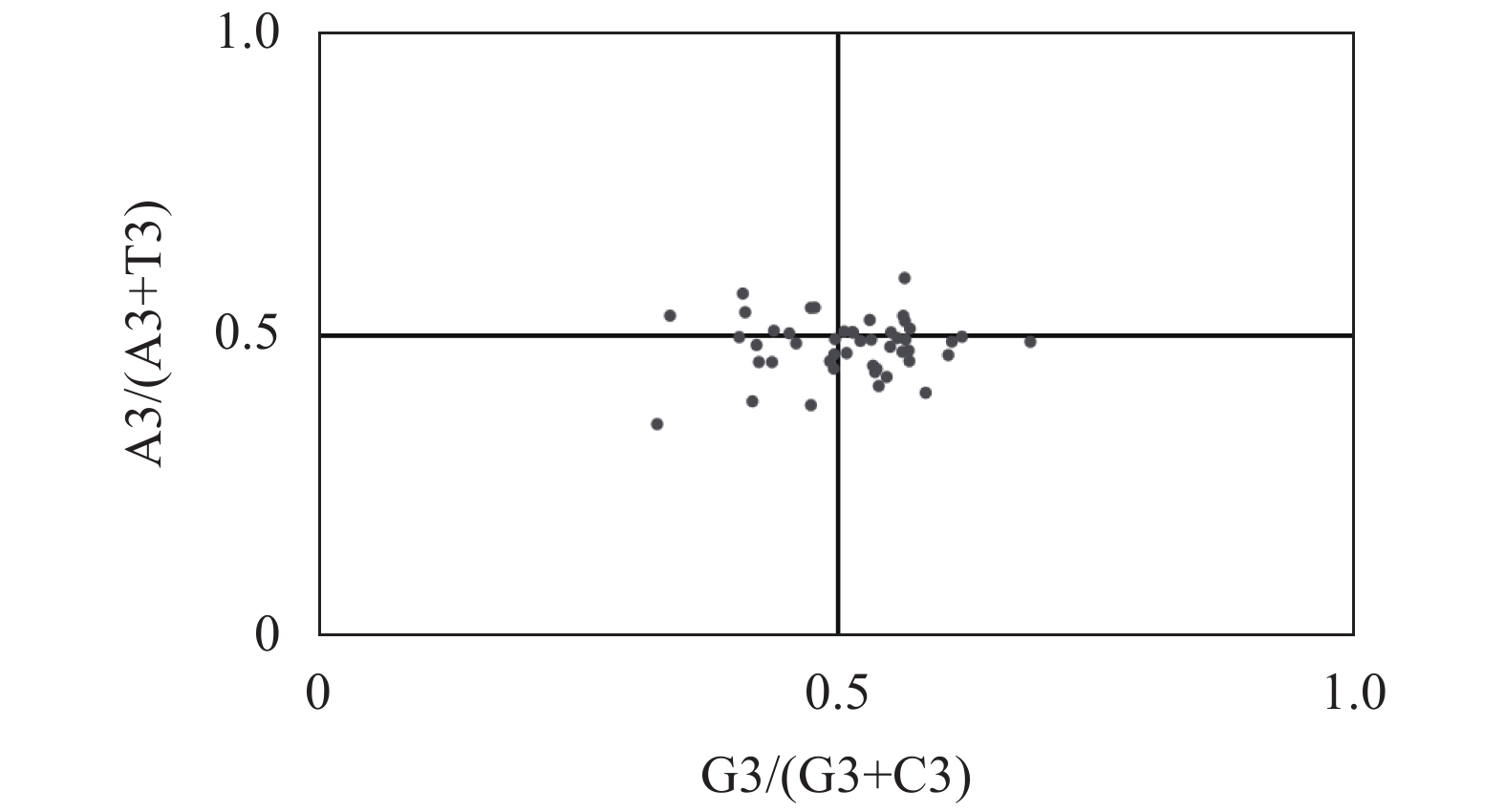
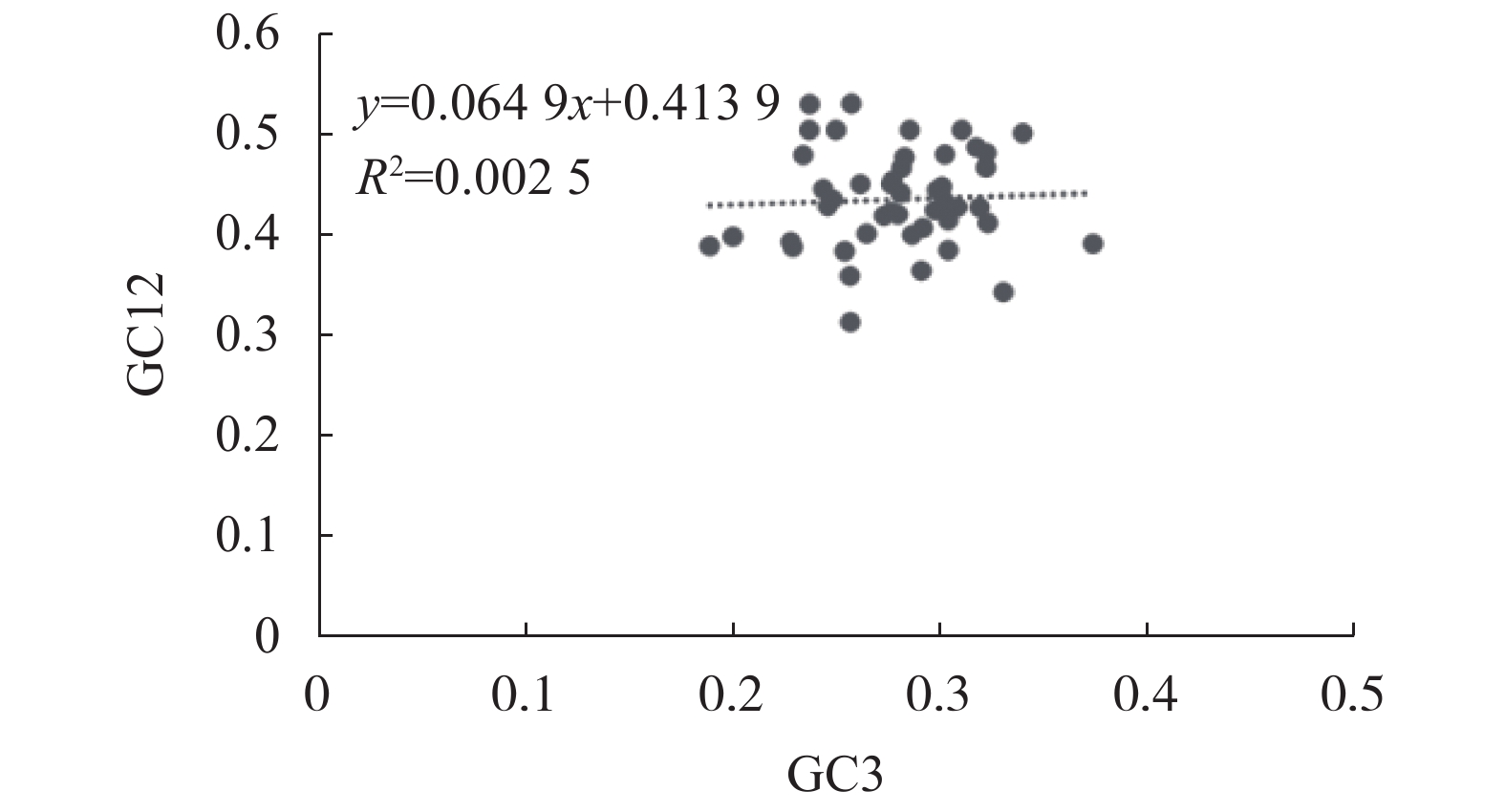
摘要:目的 分析镰翅羊耳蒜叶绿体基因组密码子偏好性,并探讨影响密码子偏好性形成的主要因素。方法 从镰翅羊耳蒜叶绿体基因组中筛选出47条编码序列,通过CodonW和CUSP软件计算不同基因的GC含量,分析密码子使用模式。结果 GC3含量为27.99%,低于GC1(47.00%)和GC2(39.41%);ENC取值范围为40.85~56.80,平均值为47.27;ENC与GC1、GC2之间均无显著相关,与GC3呈极显著正相关;GC12与GC3的相关性不显著,相关系数为0.05;47%基因的ENC比值频数分布在−0.05~0.05区间,53%基因的ENC比值频数分布在−0.05~0.05区间之外;碱基的使用频率为A>T、G>C;17个密码子被确定为最优密码子。结论 镰翅羊耳蒜叶绿体基因组密码子对A或T碱基结尾有偏好性;密码子偏好性较弱;密码子偏好性主要受选择和突变作用共同影响,同时还受其他因素影响,是多因素综合影响的结果。Abstract:Objective The coden usage bias and its infulence factor in the chloroplast genome of Liparis bootanensis were analyzed in this study.Method CodonW and CUSP software were used to analyze the codon usage bias based on 47 codon DNA sequences of the chloroplast genome of L. bootanensis.Result The GC content in the 3rd position of the codons was 27.99%, which was lower than those in the 1st and the 2nd. The effective number of codons ranged from 40.85 to 56.80, averaging 47.27. The ENC showed no significant correlation to GC1 and GC2 but a significant positive correlation to GC3. The correlation between GC12 and GC3 was not significant with a coefficient of 0.05. The frequency of ENC ratio of 47% genes was distributed between −0.05 and 0.05, while that of 53% genes outside -0.05 and 0.05 range. A PR2-plot analysis indicated the base usage frequencies to be A>T and G>C. There were 17 codons selected after screening.Conclusion The 3rd position of codons was rich in A or T. The codon usage bias was weak, complex, and affected by mutation and selection besides other factors.
-
Keywords:
- Liparis bootanensis /
- chloroplast genome /
- codon usage bias
-
0. 引言
【研究意义】乡村是土地、人口、聚落等资源组合产生的具有自然、社会、经济特征的地域综合体,与城镇互促互进、共生共存,共同构成人类活动的主要空间。从人地关系地域系统角度看,乡村地域主要有三大功能:一是提供充足的资源品(农业生产空间);二是提供环境负熵流,容纳消解污染物(生态空间);三是提供理想栖居空间(聚落空间)[1]。改革开放以来,随着社会经济和城镇化的快速发展,由于不同地区和村庄自然资源的差异性、经济发展的不均衡性和社会需求的多样性,乡村空间结构和功能发生了调整[2−3]。乡村功能类型和功能价值与需求潜力存在巨大差距,尤其是受城市化进程影响较大的区域,乡村生产、生活和生态条件差距比较明显,其功能配置也存在显著差异[4]。《乡村振兴战略规划(2018-2022年)》提出,坚持乡村振兴和新型城镇化双轮驱动,统筹城乡国土空间开发格局,优化乡村生产生活生态空间,分类推进乡村振兴,打造各具特色的现代版“富春山居图”。如何优化乡村生产生活生态空间,分类推进乡村振兴成为困扰学者们的难点。乡村衰落,直接引发人、土地和聚落的空心化。从乡村空间结构和经济结构重构的动力机制出发,将农村空间、产业和组织结合,探索乡村重构的模式和道路,是新时期乡村空间重组的核心[5]。【前人研究进展】乡村发展水平综合评判是乡村发展转型类型划分的基础[6],乡村主导功能则是乡村转型发展类型划分的重要依据。长期以来,为了更深入地认识乡村和推进乡村发展,国内外学者们对乡村发展从评价尺度、评价方法、功能理论与发展内容等进行了大量的研究。在评价尺度上,大部分的学者主要从县域宏观尺度对乡村发展进行评价。崔悦怡等[7]以西部地区各省(自治区、直辖市)为尺度评价农村综合发展情况;贾晋等[8]、闫周府等[9]分别从省域层面测度和比较了乡村振兴发展水平;张挺等[10]对11省(市)的35个乡村进行了实证评价分析;韩磊等[11]从全国、区域、省级三个层面对农村发展进程进行测度和比较研究。在评价方法上,主要有数学模型等定量评价方法和GIS与RS的空间分析方法等。刘玉等[12]运用自组织映射神经网络法(SOFM)数学模型开展环渤海地区乡村地域功能分区;洪惠坤等[13]运用GIS技术和Dagum基尼系数法,对重庆市乡村空间功能值的空间非均衡及分布动态进行了实证研究;唐林楠等[14]借助BP神经网络模型揭示北京平谷区“生产-生活-生态”功能分异特征并运用Ward法划分乡村功能理论与发展内容上,Banski等[15]根据用地特征、社会服务、就业结构等指标,把波兰苏瓦乌基省划分为以农业为主导功能,以及以工业、旅游业、林业等为优势功能的6个功能类型;Willemen等[16]在把荷兰乡村地域划分为居住、文化等功能的基础上研究了乡村地域功能间的相互作用机制;Holmes等[17]提出多功能乡村转型理论,指出社会发展过程中人们对乡村生产、消费和生态的需求变化是驱使乡村功能不断转型的主要原因;杨忍等[18]基于国土空间“生产-生活-生态”三生功能理论,把乡村功能类型划分为农业生产功能、人居生活功能和生态保育功能等8种类型;刘彥随等[19]构建地域多功能性评价指标体系与指数分析模型,对中国县域经济发展、粮食生产、社会稳定、生态保育功能及其综合功能进行分级研究;刘自强等[20]分析了不同工业化发展阶段城市地域功能与乡村地域功能的演化轨迹,并以此把乡村发展阶段划分为维持生计型、产业驱动型与多功能主导型3个阶段;李平星等[21]以经济发达的江苏省为例,采用价值评价法核算1990—2010年乡村地域生态保育、农业生产、工业发展、社会保障功能及其综合价值,分析时间演变与空间分异规律。【本研究切入点】虽然国内外学者对乡村发展的研究成果辉煌,但在研究角度取向方面尚待进一步细化。一是大部分研究成果都是针对县域以上宏观层面,从行政村特别是连片村庄进行分析的更少。由于乡村的动态性、多样性、不整合性及相对性等因素[22],乡村评价受尺度影响显著,其尺度越小,越有助于科学揭示乡村发展的本质特征,越有助于深入揭示乡村特性与乡村发展之间的空间、经济和社会关联,因此乡村评价研究呈现出微观化和应用化趋势[23]。县域以上尺度的宏观评价分析经济发展的“人” “地”矛盾,对一定地理环境乡村振兴的乡村“病”药方就难以准确。村域尺度的乡村发展评价是对乡村“病”的直接“把脉”,是村庄乡村振兴定位和乡村发展类别判断的重要依据。二是鲜见涉及城市郊区的村域“三生”功能评价。城市近郊区是城乡融合的关键地带,识别该地区村庄功能与类型有助于统筹城乡发展格局[24]。【拟解决的关键问题】县(市)是中国经济社会功能比较完整的行政地域单元,行政村是农村经济社会活动与规划管理的基本单元,在一个县(市区)从村域进行乡村发展评价研究,能更深入揭示乡村发展特征及其存在的问题。为此本文以位于福州市城郊的晋安区北部59个村庄为例,将乡村发展要素分成生产、生活和生态三个方面,构建村级乡村发展评价体系,深入分析乡村发展演变的要素差异和空间特征,并通过乡村发展聚类特征状况,以“主导空间+辅助空间”的定位分析乡村振兴的空间开发格局,以期为形成因地制宜、循序渐进的乡村振兴策略,打造各具特色的现代“富春山居图”提供方法和思路。
1. 研究方法与数据来源
1.1 指标体系构建
综合考虑评价单元的完整性、评价方法的可操作性、数据资料的可获得性以及评价指标的稳定性,建立村域尺度乡村性评价指标体系,要素层包括生产、生活、生态三大功能,指标层包含25个指标(表1)。
表 1 “三生”功能乡村发展评价指标体系Table 1. Evaluation index for rural development of “Production-living-ecological” function要素层 Element 指标层 Index 计算方法 Description 权重 Weight 生产功能
Production function(PF)粮豆人均产量/t 村粮豆总产量/总人口数 1/9 蔬果人均产量/t 村蔬果总产量/总人口数 1/9 本村就业比例/% 留守就业人数/总人口数 1/9 产值人均数/元 村生产总值/总人口数 1/9 技术人口比例/% 村技能型人员数量/总人口数 1/9 合作社数量/个 村建立并实际运营的合作社数量 1/9 村外援人才状况 3年内平均村干部挂职和下派干部人数 1/9 乡贤情况 处科级以上领导+村人才学历(博士、硕士) 1/9 通村公路硬化率/% 村内公路硬化总里程/村内公路总里程 1/9 生活功能
Living function(LF)现代房屋户比例/% 1990年后建设且完好的房屋数量/总房屋数量 1/11 活动中心情况/个 各自然村活动中心数量和 1/11 学校教育方便性/个 幼儿园数量×0.5+小学数量×0.5 1/11 生活用品采购便利性/个 村内小卖部数量和 1/11 居民文化提升机会 图书角数量×0.5+(会堂或讲堂数量)×0.5 1/11 居民办事方便状况 村中有无居民服务中心 1/11 党员比例/% 村党员人数/总人口数 1/11 党员数量/人 村党员人数 1/11 人均纯收入/元 家庭纯收入总量和/村内总人口数 1/11 农村自来水状况/% 各自然村自来水开通比例 1/11 村卫生服务状况 村卫生服务站数量 1/11 生态功能
Ecological function(EF)森林覆盖率/% 森林总面积/村地域总面积 1/5 垃圾集中处理状况/% 自然村集中垃圾处理点覆盖比例 1/5 污水处理情况/% 各主要河流定期整治数/总河流数量 1/5 美丽乡村建设状况/分 按级别设定分值(国家级5、省级4、市级3、县级1) 1/5 村规民约制定状况 有无制定村规民约及针对性比例 1/5 (1)生产功能(Production function, PF)。该功能主要从农村产业发展的角度,以外部环境条件和内部发展能力两个方面构建指标。选取包括粮豆人均产量、蔬果人均产量、本村就业比例、人均产值、技术人口比例、合作社数量、村外援人才状况、乡贤情况和通村公路硬化率9个指标。
(2)生活功能(Living function, LF)。该功能主要反映村民生活基本功能和新时期现代生活提升需求,从硬件配套、经济水平和组织能力等方面综合构建。选取包括现代房屋户比例、活动中心、学校教育方便性、生活用品采购方便性、居民文化提升机会、居民办事方便状况、党员比例、党员数量、人均纯收入、农村自来水状况和村卫生服务状况11个评价指标。
(3)生态功能(Ecological function, EF)。该功能主要从自然环境条件和公共建设等角度,按照可持续发展的原则,构建包括森林覆盖率、垃圾集中处理状况、污水集中处理状况、美丽乡村建设状况和村规民约制定状况5个评价指标。
1.2 评价模型与指标权重
采用改进后的TOPSIS评价法[25]对区域内村庄的演变进行综合评价。首先整理调查数据,构建59行25列的原始数据矩阵,对其进行归一化处理使其无量纲化,然后找出方案中的最优方案和最劣方案,分别计算各评价对象与最优方案和最劣方案间的距离,获得各评价对象与最优方案的相对接近程度,以此作为评价优劣的依据。由于指标间的重要性判断带有主观因素,并且不同主体的看法也有差别,为了减少这方面的影响,指标间的权重分配采用平均法(表1)。
评价的主要步骤如下:
(1)同趋势化。综合评价中,有些指标是高优指标,有些指标是低势指标。用TOPSIS法进行评价时,要求所有指标方向一致。通常采用将低优指标高优化,对绝对数指标使用倒数法(
1/X ),对相对数低优指标使用差值法(1−X )。(2)指标无量纲化。为了消除指标计量单位的影响,要对指标实测值进行归一化处理,即无量纲化。设
(Xij)n×m 为同趋势化后的指标矩阵,(Zij)n×m 为归一化后的数据矩阵,则:Zij=Xij/√m∑i=1X2ij ,式中j=1,2,⋯,m 。(3)确定优先方案中的最优方案
Z+ 和最劣方案Z - 。若原始数据经同趋势化统一为高优指标,则:最优方案为:
Z+=(Z+1,Z+2,⋯,Z+m) 最劣方案为:
Z-=(Z-1,Z-2,⋯,Z-m) 其中,
Z+j=max ,Z_j^{\text{ - }} = \mathop {\min }\limits_{1 \leqslant i \leqslant n} \left\{ {{Z_{ij}}} \right\} ,式中j = 1,2, \cdots ,m 。(4)计算各评价对象与最优方案和最劣方案的加权欧氏距离
D_i^ + 和D_i^{{ - }} :D_i^ + = \sqrt {\sum\limits_{j = 1}^m {{{[{W_j}({Z_{ij}} - Z_{ij}^ + )]}^2}} } D_i^{\text{ - }} = \sqrt {\sum\limits_{j = 1}^m {{{[{W_j}({Z_{ij}} - Z_{ij}^{{ - }})]}^2}} } 式中
{W_j} 为各指标权重,i = 1,2, \cdots ,n 。(5)计算各评价对象与最优方案的接近程度
{C_i} :{C_i} = \frac{{D_i^ - }}{{D_i^ + + D_i^ - }} ,式中i = 1,2, \cdots ,n 。{C_i} \in [0,1] ,{C_i} 愈接近于1,表示第i 个评价对象越接近于最优水平;反之,{C_i} 愈接近于0,表示第i 个评价对象越接近于最劣水平。即{C_i} 值越大,评价结果越优。1.3 实证对象与数据来源
本研究实证对象为位于省会城市福州郊区的晋安区,该区包括3个街道6个乡镇,其中3个街道和3个乡镇已经基本城市化。全区常住人口85.5万人,其中农业人口约7万人,主要分布于宦溪、寿山和日溪3个乡镇。选取具有农村特征的宦溪、寿山和日溪3个乡镇59个村为样本。数据来源,一是社会经济数据来自《福州市晋安区统计年鉴》(2015—2018年),土地使用数据和空间地图来自于国土资源部门;二是通过发放调查问卷的方式获取,问卷填写对象主要为村书记和村主任,总共发放问卷59份,有效问卷59份。对于问卷中存在不清楚或未填写的问题,采取电话询问加以补充。
1.4 研究思路与方法
1.4.1 研究思路
随着我国经济发展水平的提高,城乡收入水平和公共服务供给能力的差异逐渐被拉大,乡村振兴要缩小这种差距,就要在经济活动中解决“人”和“地”的匹配问题。通过聚类分析方法剖析所有村庄的类型结构,同时结合村庄空间定位,围绕中心村形成“点—线—面”区域多村结合、联村共振的发展路径,促进农村产业发展和公共服务配套优化,特别是在农村一二三产融合发展模式下,更能促进乡村的振兴,从而逐步形成产权改革机制下的产业振兴、保留乡愁特色下的文化振兴、党建引领下的组织振兴、多主体共建下的人才振兴和自然谐发展下的生态振兴,形成板块式、连片化发展。
1.4.2 分类评价
以最显著要素为主导空间,其他要素为辅助空间,构造“主导空间+辅助空间=乡村类型”的乡村定位,将乡村分成“生活+生产、生活+生态、生活+生产+生态、生产+生活、生产+生态、生产+生活+生态、生态+生活、生态+生产、生态+生活+生产”9个类型(图1)。同时,将9个类型融入集聚提升、城镇转化、特色保护、搬迁撤并的村庄类型体系中,其中集聚提升包括“生活+生产、生产+生活、生活+生产+生态”、融入城镇包括“生活+生态”、特色保护包括“生产+生活+生态、生态+生活、生态+生活+生态”、搬迁撤并包括“生态+生态、生态+生产”,为乡村重构提供融合基础。
![]() 图 1 乡村演化与乡村重构的逻辑与路径Figure 1. Logic and path of rural evolution and rural reconstruction
图 1 乡村演化与乡村重构的逻辑与路径Figure 1. Logic and path of rural evolution and rural reconstruction2. 结果与分析
2.1 评价结果
根据TOPSIS方法,得到晋安区北部山区各行政村乡村发展生产、生活和生态指数(图2),59个村的乡村发展总指数为0.18~0.48,表明晋安区北部村庄受省会城市对乡村资源吸纳影响,乡村弱化比较严重。从3个要素指标分析,生态指标最好,其次是生活指标,最后是生产指标。另外,3个要素的变异系数都比较大,全部大于20,生产要素的变异系数高达34.50,显示村之间发展不平衡。
![]() 图 2 不同行政村乡村发展指数变化Figure 2. Changes in rural development index of different administrative villages
图 2 不同行政村乡村发展指数变化Figure 2. Changes in rural development index of different administrative villages2.2 空间特征分析
图3a反映了晋安区北部59个村总体指数情况和要素指数情况,为此进一步分析总指数和要素指数在地域空间的布局状况。
![]() 图 3 乡村发展指数空间分布白色区域为市区。Figure 3. Spatial distribution of rural development indexThe white area represents urban areas.
图 3 乡村发展指数空间分布白色区域为市区。Figure 3. Spatial distribution of rural development indexThe white area represents urban areas.2.2.1 总指数的空间特征分析(RRI)
晋安区北部乡村发展评价指数(RRI)表现为北高南低的空间分布特征(图3a)。其中,低值区(RRI≤0.25)主要分布在宦溪镇镇政府所在地周边村庄和工业化开发比较大的村庄及一些交通不便的村庄,前两种村庄经济基础比较好、交通比较方便,城镇化水平比较高,第一产业比重低,后一种村庄除了生态不错,产业、区位和交通条件都比较弱。偏低值区(0.25<RRI≤0.35)为北峰山区村庄的主要部分,有34个村庄。区位和交通相对不错,主要分布于交通干线周边,资源要素比较丰富。高值区(RRI>0.35)主要是特色村庄,产业基础较好、交通条件优越、资源比较丰富且有特色。图3a显示,低值区和高值区的聚集程度不高,比较分散。而偏低值区的集聚程度比较高,特别是北部沿省道周边的村庄。从乡镇看,乡村发展指数为日溪乡(0.33)>寿山乡(0.29)>宦溪镇(0.25)。
2.2.2 生产要素的空间特征分析(PRI)
晋安区北部山区生产要素整体水平(PRI=0.26)低于总体发展水平(RRI=0.31),介于0.09~0.48,呈现中间低、南北两端高且发散趋势,空间分布差异显著(图3b),变异系数34.50。低值区(PRI≤0.25)包括27个(45.76%)行政村,这些村庄生产发展比较不足,客观原因是晋安区北部山区交通和自然条件相对不足,特别是因山区地貌限制交通受到很大制约;主观原因为晋安区位于省会城市郊区,改革开放后城市重点发展偏向于近郊的平原区,对远郊的山区发展关注不够。偏低值区(0.25<PRI≤0.35)有20个(33.90%)行政村。高值区(PRI>0.35)仅有12个(20.34%)行政村。乡镇生产要素水平排序为:日溪乡>寿山乡>宦溪镇。
2.2.3 生活要素的空间特征分析(LRI)
晋安区北部山区生活要素整体水平(LRI=0.34)高于总体发展水平(RRI=0.31),介于0.14~0.63,呈发散分布,空间分布差异显著(图3c),变异系数27.92。从指数分布区间看,低值区(LRI≤0.25)仅包括4个(45.76%)行政村,偏低值区(0.25<LRI≤0.35)有40个(33.90%)行政村,高值区(LRI>0.35)有15个(20.34%)行政村。量化分析结果,晋安区北部山区在城郊经济的带动下人均收入较高,其中务工收入有很大的贡献。在生活配套等方面,伴随美丽乡村建设,自来水入户、公路硬化、卫生服务等公共设施基本到位。但受不方便的交通条件、少而分散的耕地,以及陡峭的地形地貌,3个山区乡镇的人口仅有3.15万人,仅占晋安区总人口的6.76%,59个村户籍人口超过1000人的仅有3个,500人以下的有28个,随着外出人口的不断增加,村庄的常住人口不断减少,导致大部分的公共资源配套利用率低且逐步弱化,特别是在村级幼儿园和小学等教育资源配套方面具有很大的缺失。乡镇生活要素水平排序为:寿山乡>日溪乡>宦溪镇,其中日溪和寿山比较接近。
2.2.4 生态要素的空间特征分析(ERI)
北部山区生态要素整体水平(ERI=0.37)高于总体发展水平(RRI=0.31),介于0.17~0.66,呈东北高、西南低的趋势,空间分布差异相对较小(图3d),变异系数24.64。从指数分布区间看,低值区(ERI≤0.25)仅包括5个(45.76%)行政村,偏低值区(0.25<ERI≤0.35)有21个(33.90%)行政村,高值区(ERI>0.35)有33个(20.34%)行政村。从以上分析看,生态要素水平在3个要素指标中水平最佳。主要因素:一是晋安区北部山区的自然环境条件比较好,特别是森林覆盖率总体较高;二是通过美丽乡村建设的改造,不断提升环境整治、垃圾处理能力,生态环境持续优化。乡镇生态要素水平排序为:日溪乡>寿山乡>宦溪镇。
2.3 乡村多元发展类型分析
通过微观尺度的乡村发展评价结果(图3),可以得出59个村在生产、生活和生态的空间特征及其内部差异。通过(图1)的乡村定位方法,59个村的功能和类型如表2所示。
表 2 乡村功能与类型识别Table 2. Rural function and type identification主导空间
Dominant condition辅助空间
Auxiliary condition村庄类型
Village type村庄 Village 生活 Living 生产 集聚提升 岭头村、中心村、胜利村、前洋村 生态 融入城镇 宦溪村、日溪村、湖山村、垄头村、湖中村 生产+生态 集聚提升 红寮村、宜夏村、创新村、鹅鼻村、峨嵋村、寿山村、汶洋村、井后村、梓山村、降虎村、上寮村 生产 Production 生活 集聚提升 过仑村、江南竹村 生态 搬迁撤并 洲洋村、建立村 生活+生态 特色保护 叶洋村、点洋村、弥高村、恩顶茶场、山头顶村、 生态 Ecology 生活 特色保护 板桥村、石牌村、九峰村、芙蓉村、亥由村、溪下村、贵洋村、菜岭村、南洋村、山溪村 生产 搬迁撤并 上仑村、优山村、长基村、东坪村、增楼村、牛项村、黄土岗村、吾洋村、黄田村、
红庙村、沙溪村、大坂村、汶石村、南峰村、党洋村、铁坑村、万洋村、山秀园村生活+生产 特色保护 民义村、芹石村 2.3.1 以生活为主导的乡村发展类型
该类乡村发展有3种模式,分别是“生活+生产”“生活+生态”和“生活+(生产+生态)”。晋安区北部山区以生活为主导空间的行政村有20个。该类型村庄的特征主要表现为交通比较方便、人口相对密集、资源比较丰富。因此应结合空间邻近型乡村推进城乡空间一体化发展,破解城乡二元结构壁垒,充分利用公共服务资源和区域优势,大力发展乡村空间经济。从国家村庄类型划分看,这20个村庄可以分成集聚提升和融入城镇两类。其中集聚提升类村庄要在原有规模基础上改造提升、优化环境、保护保留乡村风貌,同时激活产业、提振人气、增添活力,建设宜居宜业的现代化村庄。融入城镇类村庄要做好基础设施互联互通、公共服务共建共享,在形态上保留乡村风貌,在景观配置上突出乡村特色,在治理上主动承接城市功能外溢,为城乡融合发展提供实践经验。
2.3.2 以生产为主导的乡村发展类型
该类乡村发展有3种模式,分别是“生产+生活”“生产+生态”和“生产+(生活+生态)”。晋安区北部山区以生产为主导空间的行政村有9个。该类型村庄的特征主要表现为农业资源和土地利用比较丰富,交通尚可,但生活条件相对滞后。因此乡村应通过资源市场化开发,积极培育优势产业,同时积极推进乡村空间重构,有效整合乡村产业、人口、土地、基础设施等要素,提高乡村经济发展活力,拓宽乡村居民的收入来源。从国家村庄类型划分看,主要是集聚提升、特色保护和搬迁撤并三类。其中集聚提升类村庄在要激活原有产业规模基础,发挥自身比较优势,强化主导产业支撑,走专业化村庄发展路径。特色保护类村庄要合理利用特色资源,发展特色产业和乡村旅游;同时要尊重原住居民生活形态和传统习惯,适当改善村庄基础设施和公共环境,形成特色资源保护与村庄发展的良性互促机制。搬迁撤并类主要为人口数量太少导致公共资源配置不足的村庄,可与就近中心村联结,通过农村集聚发展搬迁、易地搬迁、生态宜居搬迁等方式,统筹解决村民生计和优化区域生态保护等问题。
2.3.3 以生态为主导的乡村发展类型
该类乡村发展有3种模式,分别是“生态+生活”“生态+生产”和“生态+(生产+生活)”。晋安区北部山区以生态为主导空间的行政村有30个。该类型村庄的特征主要表现为资源比较丰富,但交通可达性不足、生活环境条件较差。因此该类型乡村主要在保护特色资源的前提下继续加强乡村产业特色化发展,如休闲度假区、观光农业区、生态旅游区等,不断提升乡村竞争力。对于条件比较脆弱的村庄,以环境保护为主,减少或限制开发。从国家村庄类型划分看,主要是特色保护和搬迁撤并两类。特色保护类村庄要尊重原住居民生活形态和传统习惯,促进村庄特色资源合理利用与生态保护相和谐,形成特色资源保护与村庄发展的良性互促机制。搬迁撤并类村庄要严格限制新建、扩建活动,可通过集聚发展搬迁、易地搬迁、生态宜居搬迁等方式,实施与就近中心村搬迁撤并,搬迁撤并后的村庄原址,因地制宜复垦或还绿,增加乡村生产生态空间。
2.4 村庄聚类分析与板块化发展识别
2.4.1 村庄聚类分析
根据村庄聚类结果,得到4个类别村庄(表3)。其中第一类村庄有9个,该类村庄基本属于条件比较突出,位置比较中心;第二类村庄有6个,第三类村庄有6个,这些村庄属于发展比较有优势或特色比较鲜明的村庄,村庄基本配套比较充足;第四类村庄有38个,该类村庄属于条件比较一般、资源和特色不是太突出。
表 3 聚类分析归类Table 3. Cluster analysis类型 Type 村庄 Village 一类 山溪村、红寮村、上寮村、中心村、垄头村、岭头村、宜夏村、宦溪村、日溪村 二类 鹅鼻村、石牌村、过仑村、降虎村、板桥村、黄土岗村 三类 叶洋村、上仑村、胜利村、长基村、优山村、创新村 四类 恩顶茶场、南洋村、增楼村、洲洋村、建立村、民义村、亥由村、黄田村、牛项村、弥高村、峨嵋村、湖山村、湖中村、溪下村、
前洋村、山头顶村、江南竹村、红庙村、沙溪村、吾洋村、贵洋村、菜岭村、九峰村、大坂村、芙蓉村、芹石村、寿山村、点洋村、
汶洋村、汶石村、南峰村、东坪村、党洋村、铁坑村、万洋村、井后村、梓山村、山秀园村2.4.2 村庄板块化发展识别
通过上述59个村的空间定位和聚类分析,按照“联村共振”区域板块化合并方法,晋安区北部山区59个村庄划分为12个板块(图4)。12个板块中,岭头板块、宦溪板块和捷坂板块包含的村庄达到8个,汶洋板块、党洋板块和日溪板块包含的村仅有3个。其中除了吾洋板块没有特色村,其他板块均有。
3. 结论与讨论
3.1 结论
本研究从村域微观尺度构建了乡村发展评价模型,对晋安区北部59个村从生产、生活和生态等要素进行分析与评价,把脉乡村;在评价的基础上根据要素显著情况进行空间定位,寻找“病根”;最后,通过聚类分析寻找中心村,结合村庄空间定位,形成以中心村为依托,“点—线—面”区域多村结合、联村共振的乡村振兴路径。结果如下:
(1)晋安区北部农村发展水平总体不高且不平衡,各要素发展水平空间差异明显。总体来看,要素间生产方面表现相当不足,生态环境和生活水平相对比较良好;要素内部发展水平较低的村庄主要分布在晋安区东南部的宦溪镇,西北面的寿山乡和日溪乡的村庄发展水平相对较高。
(2)由于地形地貌的限制,仅有少量分布在区域干道附近或地貌比较平坦的村庄,生活条件和产业发展较显著。其中生活显著的行政村有20个,这些村庄表现为交通比较方便、人口相对密集、资源比较丰富,可以集聚提升和融入城镇;生产显著的行政村有9个,这些村庄表现为农业资源和土地利用比较丰富,交通尚可、但生活条件比较不好,部分可以集聚提升和进行特色保护,部分搬迁撤并、消化生活功能;生态显著的行政村有30个,这些村庄表现为自然资源比较丰富,但交通可达性不足、生活环境条件较差,少量进行特色保护,大部分进行搬迁撤并。
(3)聚类分析后,晋安区中心村和生活显著的村庄耦合性高,结合交通、区位等条件,可以形成12个联村共振板块,形成板块式、连片化发展。在这些板块中,南部的板块由于村庄比较集中,形成的集聚板块村庄比较多。而远郊的日溪乡,形成的集聚板块村庄比较少,基本在3个村左右。
3.2 政策建议
我国是一个农业大国,乡村是我国的重要组成部分,新中国成立后,我国经历了工业化进程,乡村为工业和城市的发展做出了巨大贡献,但我国乡村面临着凋敝和衰落的客观事实。从2003年以来连续15年的中央一号文件都聚焦于“三农”问题,党的十七大和十八大分别提出了城乡统筹和城乡一体化的发展战略,党的十九大提出了乡村振兴战略,党的二十大作出了全面推进乡村振兴的战略部署,体现了党和政府对“三农”的高度重视。我国幅员辽阔,东西南北差异巨大,如何根据地区特点和特色,统筹城乡发展,激发乡村发展活力,增强乡村吸引力,解决人民日益增长的美好生活需要和不平衡不充分发展之间的矛盾,让农业成为有奔头的产业,让农民成为有吸引力的职业,让农村成为安居乐业的美丽家园,是我国区域化乡村振兴“三生”协调发展的主要问题,需要地方政府和学者们依据地方实际针对性规划设计,构建拥具有地域特色、协同互补、差异化发展的乡村振兴战略。针对城郊型乡村的“三生”协调发展,建议如下:
(1)在城乡发展关系上,必须重塑城乡关系,走城乡融合发展之路。要将乡村的生产、生活、生态等多重功能结合起来,构成具有自然、经济、社会特征的多村结合“联村共治”的人类活动空间地域综合体,达到与城镇互促互进、共生共存的发展格局。
(2)在产业经济发展上,一是利用当地资源禀赋和区位条件,因地制宜发展都市农业、循环农业、特色农业,开发智慧农业、现代设施农业等新业态模式,为城市提供丰富多元的绿色、优质农副产品;二是整合自然特色景观和乡村旅游资源,积极发展乡村休闲农业,满足市民对节假日休憩和旅游休闲的需求。
(3)在环境建设上,一是生态环境发展建设方面,必须坚持人与自然和谐共生,走乡村绿色发展之路;二是生活环境建设方面,减少简单式 “穿衣戴帽”的破坏运动,通过保留乡村原始特色,打造各具特色的现代版“富春山居图”;三是生产环境转变方面,通过新生代、专业化农民的引入和老农传统发展方式理念的再造,走健康和可持续的农产品生产。
-
表 1 编码基因密码子不同位置GC含量
Table 1 GC content in various positions of codon in chloroplast gene
基因 Gene GC1/% GC2/% GC3/% GCall/% ENC 基因 Gene GC1/% GC2/% GC3/% GCall/% ENC rps12 52.42 47.58 25.00 41.67 43.84 petA 53.89 35.51 26.17 38.53 48.43 psbA 49.44 43.22 32.20 41.62 43.05 rps18 37.25 45.1 30.39 37.58 43.24 atpA 54.72 40.35 23.43 39.50 43.05 rpl20 36.67 44.17 29.17 36.67 54.03 atpF 45.95 33.51 28.65 36.04 48.25 clpP 57.56 37.07 28.29 40.98 56.80 atpI 48.39 36.69 24.60 36.56 45.27 psbB 53.83 46.17 31.04 43.68 48.68 rps2 43.22 40.25 27.97 37.15 47.67 petB 47.69 41.2 30.09 39.66 43.21 rpoC2 45.91 37.29 27.30 36.83 48.68 petD 50.61 37.8 24.39 37.60 43.52 rpoC1 50.15 38.09 29.85 39.36 49.05 rpoA 45.00 34.71 26.47 35.39 48.64 rpoB 49.30 38.38 28.10 38.59 48.53 rps11 53.24 51.8 23.74 42.93 43.04 psbD 52.26 42.94 30.23 41.81 42.10 rps8 40.15 37.12 18.94 32.07 40.85 psbC 53.38 45.99 33.97 44.44 45.89 rpl14 53.66 35.77 27.64 39.02 43.44 rps14 43.56 46.53 27.72 39.27 41.37 rpl16 52.21 52.94 25.74 43.63 41.93 psaA 52.06 43.41 32.22 42.57 49.71 rps3 44.29 33.79 22.83 33.64 45.82 ycf3 47.34 39.05 24.85 37.08 53.43 rpl22 41.40 35.67 22.93 33.33 44.99 rps4 48.02 38.12 30.20 38.78 51.44 ycf2 42.53 35.21 37.34 38.36 53.23 ndhJ 47.17 37.74 30.82 38.57 53.38 ndhB 41.88 39.92 32.29 38.03 47.85 ndhK 41.53 42.80 30.08 38.14 50.43 rps7 53.85 46.15 23.72 41.24 47.64 ndhC 49.59 34.71 29.75 38.02 51.42 ccsA 34.37 38.08 29.10 33.85 47.69 atpE 50.37 42.22 28.15 40.25 44.72 ndhE 43.14 33.33 30.39 35.62 54.50 atpB 55.29 41.32 31.74 42.78 51.77 ndhG 40.11 36.16 25.42 33.9 45.14 rbcL 57.69 42.31 28.54 42.85 47.79 ndhA 42.03 37.09 20.05 33.06 42.54 accD 36.44 34.99 25.67 32.37 44.91 ndhH 48.22 35.79 27.66 37.23 47.50 ycf4 43.78 41.08 31.89 38.92 48.68 ycf1 35.72 26.67 25.69 29.36 46.69 cemA 41.74 26.52 33.04 33.77 47.70 平均值 AVG 47.00 39.41 27.99 38.13 47.27  下载: 导出CSV
下载: 导出CSV
表 2 编码基因密码子参数的相关性
Table 2 Correlations among individual related parameters of genes
项目 Items GC1 GC2 GC3 GCall ENC N GC1 1.000 GC2 0.390** 1.000 GC3 0.047 0.035 1.000 GCall 0.802** 0.758** 0.396** 1.000 ENC 0.022 −0.277 0.455** 0.029 N −0.126 −0.286 0.296* −0.118 0.196 1.000 注:*表示在0.05水平上显著相关;**表示在0.01水平上极显著相关。
Note: * means significant correlation at 0.05 level; ** means extremely significant correlation at 0.01 level.
下载: 导出CSV
表 3 相对同义密码子使用度分析
Table 3 RSCU analysis on protein coding region
氨基酸Amino acid 密码子 Coden 数量 Number RSCU 氨基酸 Amino acid 密码子 Coden 数量 Number RSCU Phe UUU 649 1.37 Ser UCU 410 1.71 UUC 359 0.63 UCC 231 0.98 Leu UUA 584 2.09 UCA 283 1.25 UUG 379 1.16 UCG 114 0.43 CUU 384 1.31 Pro CCU 287 1.47 CUC 127 0.38 CCC 172 0.89 CUA 259 0.74 CCA 206 1.15 CUG 126 0.32 CCG 75 0.31 AUU 770 1.48 Thr ACU 379 1.69 Ile AUC 312 0.57 ACC 154 0.68 AUA 492 0.95 ACA 289 1.16 Met AUG 432 1.00 ACG 103 0.47 Val GUU 369 1.45 Ala GCU 444 1.82 GUC 119 0.39 GCC 133 0.59 GUA 377 1.44 GCA 309 1.23 GUG 165 0.73 GCG 93 0.36 Tyr UAU 527 1.68 Cys UGU 157 1.51 UAC 130 0.32 UGC 53 0.36 Ter UAA 22 1.40 Ter UGA 11 0.70 UAG 14 0.89 Trp UGG 322 0.87 His CAU 330 1.45 Arg CGU 260 1.60 CAC 97 0.47 CGC 63 0.37 Gln CAA 528 1.59 CGA 246 1.31 CAG 162 0.41 CGG 83 0.43 Asn AAU 695 1.50 AGA 359 1.71 AAC 190 0.45 AGG 116 0.59 Lys AAA 772 1.50 Ser AGU 292 1.31 AAG 249 0.41 AGC 75 0.31 Asp GAU 638 1.62 Gly GGU 405 1.30 GAC 131 0.38 GGC 122 0.41 Glu GAA 822 1.52 GGA 499 1.68 GAG 271 0.48 GGG 206 0.62 注:Phe:苯丙氨酸,Leu:亮氨酸,Ile:异亮氨酸,Met:蛋氨酸, Val:缬氨酸,Tyr:酪氨酸,His:组氨酸,Gln:谷氨酰胺, Asn:天冬酰胺,Lys:赖氨酸,Asp:天冬氨酸,Glu:谷氨酸,Ser:丝氨酸, Pro:脯氨酸,Thr:苏氨酸,Ala:丙氨酸,Cys:半胱氨酸,Trp:色氨酸,Arg:精氨酸,Gly:甘氨酸,Ter:终止密码子。表5同。
Note: Phe: phenylalanine; Leu: leucine; Ile: isoleucine; Met: methionine; Val: valine; Tyr: tyrosine; His: histidine; Gln: glutamine; Asn: asparagine; Lys: lysine; Asp: aspartic acid; Glu: glutamic acid; Ser: serine; Pro: proline; Thr: threonine; Ala: alanine; Cys: cysteine; Trp: tryptophane; Arg: argnine; Gly: glycine; Ter: termination codon. Same for Table 5.
下载: 导出CSV
表 4 ENC比值频数分布
Table 4 Distribution of ENC ratios
组限
Class limits组中值
Class mid value组数
Frequency number组频
Frequency−0.25~−0.15 −0.20 1 0.02 −0.15~−0.05 −0.75 5 0.11 −0.05~0.05 0.00 22 0.47 0.05~0.15 0.10 19 0.40 合计 total 47 1.00
下载: 导出CSV
表 5 最优密码子分析
Table 5 Analysis on preferred codon
氨基酸
Amino acid密码子
Coden高表达基因
High expressed gene低表达基因
Low expressed geneΔRSCU 氨基酸
Amino acid密码子
Coden高表达基因
High expressed gene低表达基因
Low expressed geneΔRSCU 数量
NumberRSCU 数量
NumberRSCU 数量
NumberRSCU 数量
NumberRSCU Phe UUU* 53 1.61 25 1.46 0.15 Ser UCU*** 28 1.86 11 1.15 0.71 UUC 19 0.39 9 0.54 -0.15 UCC* 12 1.14 9 0.93 0.21 Leu UUA 47 1.70 21 1.89 -0.19 UCA 16 1.07 14 1.55 −0.48 UUG* 17 1.05 9 0.81 0.24 UCG 2 0.28 5 0.57 −0.29 CUU* 26 1.76 18 1.48 0.28 Pro CCU*** 21 1.89 10 0.91 0.98 CUC 6 0.43 5 0.37 0.06 CCC*** 10 1.03 3 0.29 0.74 CUA 24 0.84 10 0.90 −0.06 CCA 11 0.93 9 0.90 0.03 CUG 8 0.22 7 0.56 −0.34 CCG 2 0.15 3 0.29 −0.14 Ile AUU* 47 1.36 28 1.22 0.14 Thr ACU* 23 1.53 13 1.36 0.17 AUC 10 0.30 13 0.61 −0.31 ACC* 11 0.97 8 0.76 0.21 AUA* 39 1.33 26 1.16 0.17 ACA* 18 1.37 9 1.14 0.23 Met AUG 27 1.00 20 1.00 0.00 ACG 3 0.13 6 0.74 −0.61 Val GUU*** 27 2.19 17 1.55 0.64 Ala GCU 37 1.69 27 2.08 −0.39 GUC* 10 0.41 4 0.29 0.12 GCC 6 0.47 8 0.52 −0.05 GUA 20 1.15 14 1.43 −0.28 GCA*** 26 1.50 14 0.93 0.57 GUG 4 0.24 5 0.73 −0.49 GCG 6 0.34 6 0.47 −0.13 Tyr UAU* 31 1.88 30 1.67 0.21 Cys UGU 10 1.73 5 1.73 0.00 UAC 4 0.12 6 0.33 −0.21 UGC 2 0.27 2 0.27 0.00 Ter UAA 3 1.80 3 1.80 0.00 Ter UGA 1 0.60 1 0.60 0.00 UAG 1 0.60 1 0.60 0.00 Trp UGG 28 1.00 13 0.80 0.20 His CAU 15 1.50 15 1.54 −0.04 Arg CGU** 21 1.60 12 1.21 0.39 CAC 2 0.50 5 0.46 0.04 CGC 3 0.26 3 0.36 −0.10 Gln CAA 28 1.61 24 1.75 −0.14 CGA* 21 1.64 13 1.38 0.26 CAG* 7 0.39 4 0.25 0.14 CGG 1 0.08 7 0.79 −0.71 Asn AAU 27 1.50 33 1.82 −0.32 AGA** 22 1.83 15 1.50 0.33 AAC** 10 0.50 4 0.18 0.32 AGG 7 0.60 8 0.76 −0.16 Lys AAA 34 1.61 23 1.61 0.00 Ser AGU** 17 1.37 12 1.21 0.16 AAG 10 0.39 7 0.39 0.00 AGC 2 0.28 5 0.59 −0.16 Asp GAU 18 0.97 31 1.92 −0.95 Gly GGU*** 35 1.31 7 0.68 0.63 GAC* 8 0.24 2 0.08 0.16 GGC 13 0.44 8 0.60 −0.16 Glu GAA*** 44 1.76 32 1.03 0.73 GGA 29 1.73 18 1.92 −0.19 GAG 6 1.03 15 0.97 0.06 GGG 11 0.52 5 0.79 −0.27 注:_表示RSCU 值大于1 的密码子; *表示△RSCU≥0.08; **表示ΔRSCU≥0.3; ***表示ΔRSCU≥0.5。
Note: Underlining indicates RSCU>1; * means △RSCU≥0.08; ** means △RSCU≥0.3; *** means ΔRSCU≥0.5.
下载: 导出CSV
-
[1] CHEN S C, WOOD J J, ORMEROD P. Liparis L. C. Richard. In: WU Z Y, RAVEN P H, HONG D. (Eds.) Flora of China, vol. 25[M]. Science Press, Beijing & Missouri Botanical Garden Press, St. Louis, 2009: 211–228.
[2] 乔永刚, 贺嘉欣, 王勇飞, 等. 药用植物苦参的叶绿体基因组及其特征分析 [J]. 药学学报, 2019, 54(11):2106−2112. QIAO Y G, HE J X, WANG Y F, et al. Analysis of chloroplast genome and its characteristics of medicinal plant Sophora flavescens [J]. Acta Pharmaceutica Sinica, 2019, 54(11): 2106−2112.(in Chinese)
[3] DANIELL H, KHAN M S, ALLISON L. Milestones in chloroplast genetic engineering: an environmentally friendly era in biotechnology [J]. Trends in Plant Science, 2002, 7(2): 84−91. DOI: 10.1016/S1360-1385(01)02193-8
[4] LIANG W, GUO X, NAGLE D G, et al. Genus Liparis: A review of its traditional uses in China, phytochemistry and pharmacology [J]. Journal of Ethnopharmacology, 2019, 234: 154−171. DOI: 10.1016/j.jep.2019.01.021
[5] 国家中医药管理局编委会. 中华本草[M]. 上海: 上海科学技术出版社, 1999. [6] 方志先, 廖朝林. 湖北恩施药用植物志(下册)[M]. 武汉: 湖北科学技术出版社, 2006. [7] 吴润. 镰翅羊耳蒜中化学成分的研究[D]. 成都: 西南交通大学, 2017. WU R. Study on chemical constituents of Liparis bootanensis[D]. Chengdu: Southwest Jiaotong University, 2017. (in Chinese).
[8] 林爱英. 福建3种野生兰科植物繁殖生物学的初步研究[D]. 福州: 福建师范大学, 2015. LIN A Y. A preliminary study on reproductive biology of three wild orchids in Fujian province[D]. Fuzhou: Fujian Normal University, 2015. (in Chinese).
[9] LIU J F. The complete chloroplast genome sequence of Liparis bootanensis (Orchidaceae) [J]. Mitochondrial DNA Part B, 2020, 5(3): 2058−2059. DOI: 10.1080/23802359.2020.1763866
[10] BULMER M. The selection-mutation-drift theory of synonymous codon usage [J]. Genetics, 1991, 129: 897−907. DOI: 10.1093/genetics/129.3.897
[11] ROMERO H, ZAVALA A, MUSTO H. Codon usage in Chlamydia trachomatis is the result of strand-specific mutational biases and a complex pattern of selective forces [J]. Nucleic Acids Research, 2000, 28(10): 2084−2090. DOI: 10.1093/nar/28.10.2084
[12] HIRAOKA Y, KAWAMATA K, HARAGUCHI T, et al. Codon usage bias is correlated with gene expression levels in the fission yeast Schizosaccharomyces pombe [J]. Genes to Cells, 2009, 14(4): 499−509. DOI: 10.1111/j.1365-2443.2009.01284.x
[13] SHARP P M, EMERY L R, ZENG K. Forces that influence the evolution of Codon bias [J]. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 2010, 365(1544): 1203−1212. DOI: 10.1098/rstb.2009.0305
[14] 杨国锋, 苏昆龙, 赵怡然, 等. 蒺藜苜蓿叶绿体密码子偏好性分析 [J]. 草业学报, 2015, 24(12):171−179. DOI: 10.11686/cyxb2015016 YANG G F, SU K L, ZHAO Y R, et al. Analysis of Codon usage in the chloroplast genome of Medicago truncatula [J]. Acta Prataculturae Sinica, 2015, 24(12): 171−179.(in Chinese) DOI: 10.11686/cyxb2015016
[15] 王婧, 王天翼, 王罗云, 等. 沙枣叶绿体全基因组序列及其使用密码子偏性分析 [J]. 西北植物学报, 2019, 39(9):1559−1572. WANG J, WANG T Y, WANG L Y, et al. Assembling and analysis of the whole chloroplast genome sequence of Elaeagnus angustifolia and its Codon usage bias [J]. Acta Botanica Boreali-Occidentalia Sinica, 2019, 39(9): 1559−1572.(in Chinese)
[16] WRIGHT F. The ‘effective number of codons’ used in a gene [J]. Gene, 1990, 87(1): 23−29. DOI: 10.1016/0378-1119(90)90491-9
[17] JIANG Y, DENG F, WANG H L, et al. An extensive analysis on the global Codon usage pattern of baculoviruses [J]. Archives of Virology, 2008, 153(12): 2273−2282. DOI: 10.1007/s00705-008-0260-1
[18] SUEOKA N. Directional mutation pressure and neutral molecular evolution [J]. Proceedings of the National Academy of Sciences of the United States of America, 1988, 85(8): 2653−2657. DOI: 10.1073/pnas.85.8.2653
[19] SUEOKA N. Near homogeneity of PR2-bias fingerprints in the human genome and their implications in phylogenetic analyses [J]. Journal of Molecular Evolution, 2001, 53(4/5): 469−476.
[20] 胡莎莎, 罗洪, 吴琦, 等. 苦荞叶绿体基因组密码子偏爱性分析 [J]. 分子植物育种, 2016, 14(2):309−317. HU S S, LUO H, WU Q, et al. Analysis of Codon bias of chloroplast genome of Tartary buckwheat [J]. Molecular Plant Breeding, 2016, 14(2): 309−317.(in Chinese)
[21] 续晨, 贲爱玲, 蔡晓宁. 蝴蝶兰叶绿体基因组密码子使用的相关分析 [J]. 分子植物育种, 2010, 8(5):945−950. DOI: 10.3969/mpb.008.000945 XU C, BEN A L, CAI X N. Analysis of synonymous Codon usage in chloroplast genome of Phalaenopsis aphrodite subsp. Formosana [J]. Molecular Plant Breeding, 2010, 8(5): 945−950.(in Chinese) DOI: 10.3969/mpb.008.000945
[22] 李冬梅, 吕复兵, 朱根发, 等. 文心兰叶绿体基因组密码子使用的相关分析 [J]. 广东农业科学, 2012, 39(10):61−65. DOI: 10.3969/j.issn.1004-874X.2012.10.019 LI D M, LV F B, ZHU G F, et al. Analysis on Codon usage of chloroplast genome of Oncidium Gower Ramsey [J]. Guangdong Agricultural Sciences, 2012, 39(10): 61−65.(in Chinese) DOI: 10.3969/j.issn.1004-874X.2012.10.019
[23] 刘慧, 王梦醒, 岳文杰, 等. 糜子叶绿体基因组密码子使用偏性的分析 [J]. 植物科学学报, 2017, 35(3):362−371. DOI: 10.11913/PSJ.2095-0837.2017.30362 LIU H, WANG M X, YUE W J, et al. Analysis of Codon usage in the chloroplast genome of Broomcorn millet (Panicum miliaceum L.) [J]. Plant Sclence Journal, 2017, 35(3): 362−371.(in Chinese) DOI: 10.11913/PSJ.2095-0837.2017.30362
[24] INGVARSSON P K. Gene expression and protein length influence Codon usage and rates of sequence evolution in Populus tremula [J]. Molecular Biology and Evolution, 2007, 24(3): 836−844.
[25] 陆奇丰, 骆文华, 黄至欢. 两种梧桐叶绿体基因组密码子使用偏性分析 [J]. 广西植物, 2020, 40(2):173−183. DOI: 10.11931/guihaia.gxzw201811035 LU Q F, LUO W H, HUANG Z H. Codon usage bias of chloroplast genome from two species of Firmiana Marsili [J]. Guihaia, 2020, 40(2): 173−183.(in Chinese) DOI: 10.11931/guihaia.gxzw201811035
-
期刊类型引用(3)
1. 王沛然,张家林,高祥斌,涂佳才. 水肥耦合对‘聊红’椿幼苗生长特性的影响. 山东农业大学学报(自然科学版). 2024(02): 153-161 .  百度学术
百度学术
2. 王沛然,张家林,高祥斌,涂佳才. 水肥耦合对‘聊红’椿幼苗根系形态特征的影响. 山东农业科学. 2024(05): 87-95 . 百度学术
3. 尹明华,郭依瑶,李启诺,罗泽全,林文龙,熊姿雯. 信前胡烟草花叶病毒lncRNA测序鉴定、原核蛋白表达及其序列分析. 中国农学通报. 2024(21): 106-113 . 百度学术
其他类型引用(1)
































计量
- 文章访问数: 858
- HTML全文浏览量: 268
- PDF下载量: 36
- 被引次数: 4
