

Light Response Curves and Photosynthesis Models on Plants Exposed to Monochromatic Lights
-
摘要:目的 研究不同植物幼苗在多种单色光条件下的光合响应曲线及其数学模型,为植物工厂作物栽培架等现代农业种植设施的研发提供理论依据。方法 应用植物特征光谱试验方法及其测试系统,获取10种植物幼苗的22种单色光的光合响应曲线;选择红椎与马褂木两个具有代表性的植物,得出对应单色光光合响应曲线的数学模型;根据渐近线型单色光植物光合响应曲线的特点提出准饱和点光强的概念及计算方法,再依据所测定的植物幼苗22种单色光的饱和点光强,比较理论计算值与实际测定值的偏差程度,并以此为基础绘制植物光合作用饱和点曲线。结果 植物幼苗的光合响应曲线可分为两类,以红椎为代表的一类植物光合响应曲线呈渐近线型,这类曲线没有极值;以马褂木为代表的一类植物光合响应曲线则有拐点,这类曲线有极值。模型计算得到的准饱和点光强与实际测定的饱和点光强具有较高的拟合度,经过对数据的综合处理,获得对应的饱和点曲线和补偿点曲线。结论 采用植物特征光谱试验方法及其测试系统可以得到植物在单色光条件下的光合响应曲线,揭示了不同植物光合作用的规律及特点。由此提出的植物光合响应数学模型与实测值具有较高拟合度,可作为研究分析植物光合作用现象的重要参考。Abstract:Objective Mathematical models of the photosynthetic responses of plants exposed to monochromatic lights were constructed for the development of indoor agricultural cultivation.Method An experimentation was designed to generate varied wavelengths of monochromatic lights and test, measure, and construct a mathematical model for the photosynthetic responses of plants exposed to the light source. On seedlings of 10 different plant species grown under 22 varied monochromatic lights, the photosynthetic rates of the plants were measured to obtain light response curves (LRCs). Using Castanopsis hystrix and Liriodendron chinense (Hemsl.) Sarg as two representative plants, mathematical models were constructed. From the asymptotic linear curve, the light intensity at quasi-saturation point was calculated. Then, the calculated theoretical and measured empirical data were compared to test the validity of the prediction model.Result Of all the LRCs, there were two types—one of C. hystrix, which was asymptotic linear showing no plateau, and another of L. chinense, which had an inflection point. A high degree of fitting was found between the light intensity at quasi-saturation point calculated from the model and that measured empirically. After a data treatment, curves with light saturation and compensation points were obtained.Conclusion The specially designed monochromatic lights generating method and plant photosynthesis measuring system enabled a successful construction of the LRCs. The patterns and characteristics of the photosynthesis of different plants in response to the monochromatic lights varied. The high fitting on the photosynthetic response data between the values calculated from the mathematical model and those measured from the experiment suggested an applicability of the current methodology for studies on plant photosynthesis. Since the developed method and test system were not species-specific, they could be used universally for research on other plants. And using the mathematical model, optimal light wavelength and intensity at saturation point could be estimated for designing artificially controlled agriculture facilities such as “plant factories” for indoor mass crop cultivation.
-
0. 引言
【研究意义】鹿茸菇,学名荷叶离褶伞[Lyophyllum decastes (Fr.) Singer][1],是一种珍稀的药食两用型高等真菌。鹿茸菇口感爽脆、味道清淡,备受老百姓的喜爱,成为“网红菇”[2]。国内市场对于鹿茸菇的巨大需求,推动了鹿茸菇产业的快速发展,使其成为继金针菇、杏鲍菇、真姬菇、双孢蘑菇之后的新型食用菌工厂化栽培品种,生产基地主要聚集于上海、江苏、浙江、福建等地相关食用菌企业。菌丝培养阶段在整个食用菌栽培过程中十分重要。鹿茸菇菌丝在栽培基质中萌发时间长,抗杂菌能力弱[3],易发生有害菌侵害,污染率高,后期出菇产量低甚至不出菇[4]。目前鹿茸菇工厂化栽培多以固体菌种为主,传统固体菌种存在萌发时间长、制种周期长、菌龄不一致[5]、污染率高等弊端,而液体菌种培养工序简单[6],扩繁迅速,短时间内可达到一定的生物量[7-9],菌球更易分散,菌丝萌发快[10]。液体菌种较固体菌种更适用于鹿茸菇工厂化生产,因此急需开展鹿茸菇液体菌种配方优化研究。【前人研究进展】第二次世界大战期间,为了满足对抗生素的大量需求,发酵罐制造业开始兴起,同时解决了液体深层发酵的供氧问题,奠定了现代发酵工业的基础。Humfeld [11]首次使用液体深层发酵技术成功制备Agaricu campestris菌丝体。随着发酵工程理论知识与实践经验的不断积累,液体深层发酵技术现已广泛应用于大多数食用菌品种[5](除混合种出菇类型,如银耳)生产用种的制备。与固体菌种相比,液体菌种优势更明显,应用前景更广阔[12],制备工艺具备完全替代固体菌种的潜能。关于鹿茸菇方面的研究主要聚焦于鹿茸菇菌丝在平板培养基上的生物学特性,包括不同碳源、氮源、金属离子、生长素对鹿茸菇平板菌丝生长的影响[13-14],摇瓶试验分析不同培养基成分对鹿茸菇液体菌丝生长的影响[15],通气搅拌式发酵罐中优化鹿茸菇液体菌丝发酵条件,液体菌种配方优化等[16]。张汉燚等[17]对鹿茸菇中试发酵培养基进行优化,最佳培养基配方为玉米面5%、大豆0.5%、ZnSO4 0.025%、MgSO4 0.05%、KH2PO4 0.05%。发酵至第8 天时,菌丝体生物量达到最大值(10.578 g·L−1)。席亚丽等[18]对鹿茸菇摇瓶发酵条件进行探索,获得最佳培养基配方为玉米面200 g·L−1、蔗糖 15 g·L−1、麸皮35 g·L−1、酵母膏1.5 g·L−1;温度26℃,接种量10%,培养时间为6~8 d。【本研究切入点】鹿茸菇液体菌种培养优化的研究多采用单因素和正交试验结合的方式,试验设计考虑多种因素的影响,但无法明确因素之间交互作用对响应值的影响,缺少各因素与响应值之间的函数关系,无法确定最大响应值所对应的因素组合方式。目前,单因素试验和Box-Behnken响应面法结合优化鹿茸菇液体菌种发酵配方的研究较少,鹿茸菇液体发酵培养工厂化生产效率有待提高。【拟解决的关键问题】本研究以鹿茸菇为研究材料,采用摇瓶培养方式,以提高菌丝生物量为目的,采用单因素试验和Box-Behnken响应面法结合的方式,对培养基碳源、氮源和无机盐的添加量进行优化,以期获得适合鹿茸菇菌丝生长的液体发酵培养基配方,为推动鹿茸菇液体菌种生产提供理论依据。
1. 材料与方法
1.1 试验材料
1.1.1 供试菌株
鹿茸菇菌种(编号:LDS-1-YJY-2018)保存于福建农林大学(古田)菌业研究院,为福建珍菌子生物科技有限公司生产菌种。
1.1.2 试剂
酵母粉、蛋白胨均购于Oxoid;琼脂粉购于Chembase;葡萄糖、蔗糖、鱼粉、维生素B1、K2HPO4、MgSO4·7H2O、ZnSO4、CaSO4等无机盐均购于国药集团化学试剂有限公司;供试碳源玉米粉、小麦粉、全麦粉均购于陇龙之家,玉米芯购于联丰农产品深加工;供试氮源豆粕粉、牛肉膏、黄豆饼粉、花生饼粉、棉籽饼粉、玉米浆均购于鸿润宝顺培养基原料厂家。
1.1.3 培养基配方
PDA加富液体培养基(1 L):土豆200 g,葡萄糖20 g,蛋白胨2 g,酵母粉2 g,K2HPO4 2.5 g,MgSO4·7H2O 1 g,Vitamin B1 0.1 g,pH自然。PDA加富固体培养基在液体培养基的基础之上加入琼脂20 g。
摇瓶一级种子液培养基(1 L):葡萄糖20 g,蛋白胨2 g,酵母粉2 g,K2HPO4 2.5 g,MgSO4·7H2O 1 g,pH自然。
摇瓶基础发酵培养基(1 L):葡萄糖20 g,蛋白胨2 g,K2HPO4 2.5 g,MgSO4·7H2O 1 g,pH自然。
固体菌种培养基:木屑78%,麸皮20%,葡萄糖1%,石灰1%,含水率65%左右。
1.2 试验方法
1.2.1 摇瓶一级种子液的制备
将保藏菌种接种于PDA加富固体培养基平板,25℃,培养20 d左右直至菌丝长满平板。应用打孔器(0.5 mm直径)在平板上面打30孔,应用接种器将菌块挑入摇瓶一级种子液培养基(含磁力搅拌子),25℃,150 r·min−1培养4 d,第4 天取出摇瓶置于磁力搅拌器上以800 r·min−1、30 min打散菌球,继续培养4 d,即制成一级种子液。
1.2.2 液体菌种生长动力学曲线的测定
按10%接种量将一级种子液接种于100 mL基础发酵培养基,25℃、150 r·min−1振荡培养。每隔1 d分别取出发酵液,100目纱布过滤收集菌球,RO水清洗3遍,60℃烘干至恒重。每个处理设置3个重复。
1.2.3 最适液体发酵培养基成分筛选
基础发酵培养基121℃,灭菌20 min。其中玉米芯、玉米粉、小麦粉、全麦粉、豆粕粉、黄豆饼粉、花生饼粉、棉籽饼粉、鱼粉需115℃灭菌40 min,冷却后均要求过100目筛网,滤液加入基础发酵培养基。按10%接种量接入相应培养基,25℃,150 r·min−1,培养8 d,100目纱布过滤收集菌球,RO水清洗3遍,60℃烘干至恒重。每个处理设置3个重复。该操作方法适用于1.2.3、1.2.4与1.2.5。
以摇瓶基础发酵培养基为基础,分别用不同种类的碳源、氮源、无机盐替代摇瓶基础发酵培养基相对应的培养基成分,以筛选最适合鹿茸菇液体菌球生长的碳源、氮源与无机盐。供试碳源(20 g·L−1):葡萄糖、蔗糖、玉米芯、玉米粉、小麦粉、全麦粉,CK为不加碳源;供试氮源(2 g·L−1):蛋白胨、豆粕粉、牛肉膏、黄豆饼粉、花生饼粉、棉籽饼粉、鱼粉、玉米浆,CK为不加氮源;供试无机盐(1 g·L−1):CoCl2、CuSO4、ZnSO4、CaSO4、MnSO4、FeSO4、NaCl、KCl、MgSO4·7H2O、K2HPO4,CK为不加无机盐。
1.2.4 单因素设计
以摇瓶基础发酵培养基为基础,分别替换其中碳源、氮源、无机盐成分进行单因素试验。各因素质量浓度梯度如下:全麦粉为10、20、30、40、50、60、70 g·L−1;花生饼粉为2、6、10、14、18、22、26 g·L−1;K2HPO4为 0.5、1.0、1.5、2.0、2.5、3.0 g·L−1;MgSO4·7H2O为0.5、1.0、1.5、2.0、2.5、3.0 g·L−1。
1.2.5 响应面设计试验
基于单因素设计结果,以菌丝生物量为响应值评价指标,采用Box-Behnken响应面法对全麦粉(A)、花生饼粉(B)、磷酸氢二钾(K2HPO4,C)、七水硫酸镁(MgSO4·7H2O,D)进行4因素3水平优化设计试验(表1)。
表 1 Box-Behnken设计因素水平及编码Table 1. Codes and levels of factors in Box-Behnken experiment水平
Level因素 Factors/(g·L−1) A 全麦粉
Whole wheat flourB 花生饼粉
Peanut mealC 磷酸氢二钾
K2HPO4/D七水硫酸镁
MgSO4·7H2O/+1 55 24 2.25 2.25 0 50 22 2.00 2.00 −1 45 20 1.75 1.75 1.2.6 液体菌种和固体菌种出菇试验
按照最适发酵培养基配方制备液体菌种,以10%的接种量接种于250 mL摇瓶内,24℃ 130 r·min−1培养8 d,获得液体菌种。采用袋口接种的方式将液体栽培种接种至菌袋内,每袋接种25~30 mL,接种固体菌种作为对照组。养菌与出菇管理工艺均由福建珍菌子生物科技有限公司完成。
1.3 统计分析
培养基成分筛选试验和单因素试验使用Graphpad Prism 9软件作图。响应面试验使用Design Expert 8.0.6软件进行。试验数据均使用SPSS Statistics 26软件进行显著性差异分析。
2. 结果与分析
2.1 鹿茸菇液体菌种生长动力学曲线
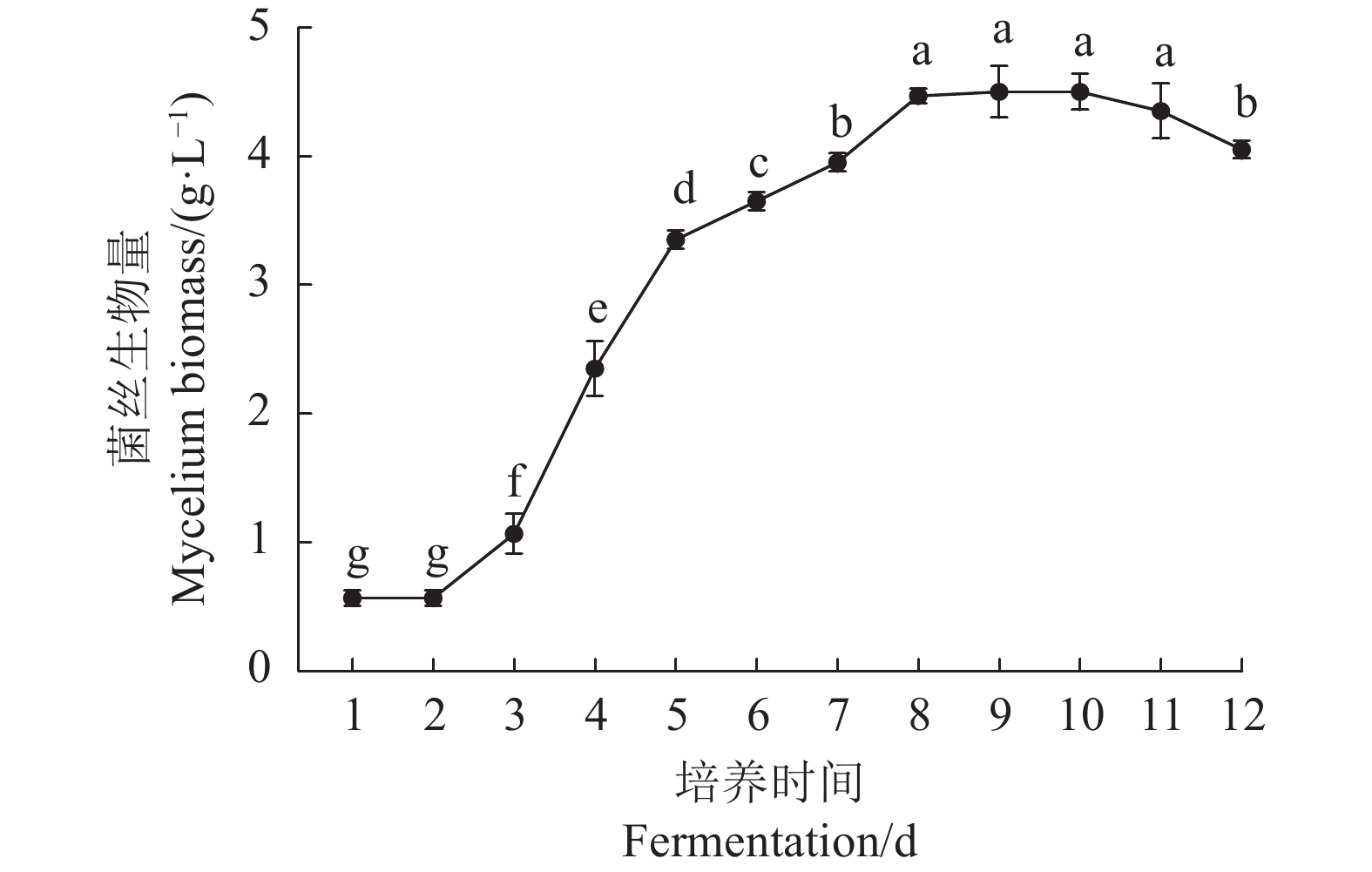
由图1可知,鹿茸菇菌丝生物量随培养时间的延长呈先上升后下降的趋势。在发酵过程中,0~2 d为延滞期,2~8 d为快速生长期,8~10 d为稳定期,10 d后进入衰亡期。当鹿茸菇菌丝培养至第8 天时,菌丝生物量达到最大值(4.47 g·L−1),因此确定鹿茸菇液体菌种培养时间为8 d。
![]() 图 1 鹿茸菇液体菌种生长动力学曲线不同字母表示均值之间差异显著(P<0.05),下同。Figure 1. The growth dynamics curve of L. decastes liquid strainData with different letters indicate significant differences at P<0.05. Same for the following figures and tables.
图 1 鹿茸菇液体菌种生长动力学曲线不同字母表示均值之间差异显著(P<0.05),下同。Figure 1. The growth dynamics curve of L. decastes liquid strainData with different letters indicate significant differences at P<0.05. Same for the following figures and tables.2.2 培养基成分的筛选
由图2-A可知,鹿茸菇菌丝在6种供试碳源中均可生长,且菌丝生物量均高于不加碳源的对照组。不同碳源对菌丝生物量的影响具有显著差异性,从高到低的顺序为:全麦粉>葡萄糖>蔗糖>小麦粉>玉米粉>玉米芯,以全麦粉作为碳源时,菌丝生物量达到最大值(8.50 g·L−1),显著高于其他碳源。由图2-B可知,鹿茸菇菌丝在8种供试氮源中均可生长,其中花生饼粉是供试氮源中最适宜鹿茸菇生长的复合氮源,以花生饼粉作为氮源时,菌丝生物量可达11.12 g·L−1。由图2-C可知,当Co2+与Cu2+质量浓度为1 g·L−1时,菌丝生物量均低于不加无机盐的对照组,抑制鹿茸菇菌丝的正常生长,而其余离子均不同程度地促进菌丝的生长发育,其中尤以K+与Mg2+最为显著。以上数据表明,较适合于鹿茸菇菌丝发酵的碳源、氮源、无机盐分别为全麦粉、花生饼粉、K2HPO4与MgSO4·7H2O。
![]() 图 2 3类营养要素对鹿茸菇液体菌丝生长的影响A、B与C分别代表6种碳源、8种氮源与10种金属离子对鹿茸菇液体菌丝生长的影响。Figure 2. Effects of 3 type nutrients on growth of L. decastes in liquid mediumA, B, and C represent the effects of 6 carbon sources, 8 nitrogen sources, and 10 metal ions on the growth of L. decastes liquid strain, respectively.
图 2 3类营养要素对鹿茸菇液体菌丝生长的影响A、B与C分别代表6种碳源、8种氮源与10种金属离子对鹿茸菇液体菌丝生长的影响。Figure 2. Effects of 3 type nutrients on growth of L. decastes in liquid mediumA, B, and C represent the effects of 6 carbon sources, 8 nitrogen sources, and 10 metal ions on the growth of L. decastes liquid strain, respectively.2.3 单因素试验
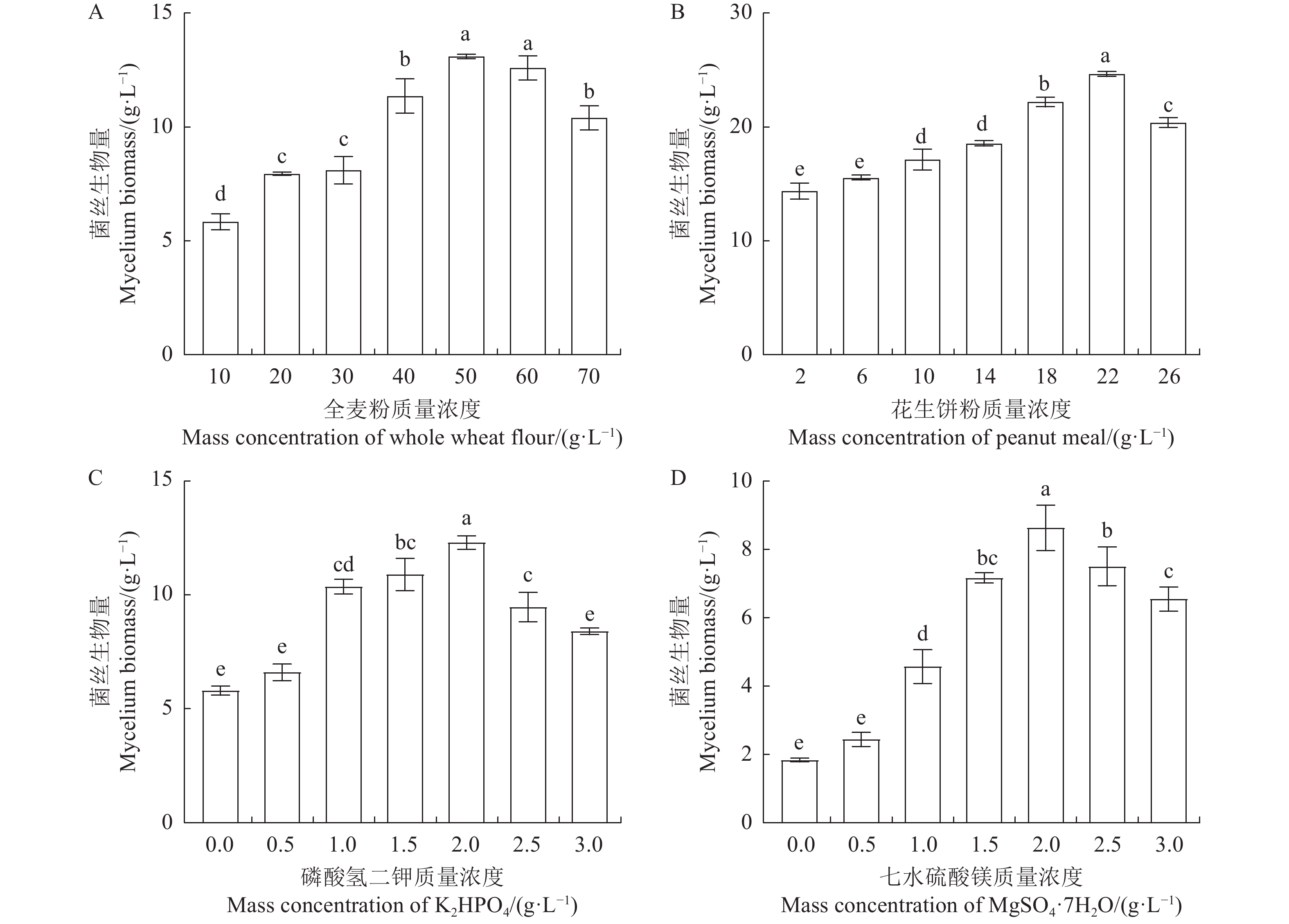
为了探究培养基单一成分如何影响鹿茸菇液体菌丝的生长,分别开展了全麦粉、花生饼粉、K2HPO4与MgSO4·7H2O单因素质量浓度梯度试验。以全麦粉作为培养基的碳源,质量浓度为40~60 g·L−1时,菌丝生物量达到最大值(13.10 g·L−1)(图3-A)。以花生饼粉作为培养基的氮源时,质量浓度为18~26 g·L−1时,菌丝生物量达到最大值(24.65 g·L−1)(图3-B)。以K2HPO4与MgSO4作为培养基的无机盐时,K2HPO4与MgSO4·7H2O的质量浓度均为1.5~2.5 g·L−1,菌丝生物量分别达到其最大值(图3-C、D)。在不考虑培养基成分之间是否存在交互作用的影响下,单因素设计已成功筛选到各培养基成分较适合鹿茸菇菌丝生长的质量浓度范围。
![]() 图 3 单因素试验A、B、C与D分别代表全麦粉、花生饼粉、K2HPO4与MgSO4·7H2O剂量梯度对鹿茸菇液体菌丝生物的影响。Figure 3. Results of single-factor design experimentA, B, C, and D: Effects of dose-dependent whole wheat flour, peanut cake flour, K2HPO4, and MgSO4·7H2O, respectively, on growth of L. decastes in liquid medium.
图 3 单因素试验A、B、C与D分别代表全麦粉、花生饼粉、K2HPO4与MgSO4·7H2O剂量梯度对鹿茸菇液体菌丝生物的影响。Figure 3. Results of single-factor design experimentA, B, C, and D: Effects of dose-dependent whole wheat flour, peanut cake flour, K2HPO4, and MgSO4·7H2O, respectively, on growth of L. decastes in liquid medium.2.4 响应面优化试验
在单因素试验结果的基础上,采用Box-Behnken设计对碳源(全麦粉)、氮源(花生饼粉)、无机盐(K2HPO4与MgSO4·7H2O)进行4因素3水平响应面优化试验,结果见表2。以因素为自变量,菌丝生物量为响应值,使用Design Expert 8.0.6软件进行多元线性回归和二次项拟合,可得到二次多项回归方程式(1),方差分析见表3。
表 2 Box-Behnken试验设计及试验结果Table 2. Design and results of Box-Behnken experiment试验号
NumberA全麦粉
Whole wheat
flourB花生饼粉
Peanut
mealC磷酸氢
二钾
K2HPO4D七水
硫酸镁
MgSO4·7H2O菌丝生物量
Mycelium
biomass/
(g·L−1)1 −1 −1 0 0 29.61±1.18 2 +1 −1 0 0 29.90±1.45 3 −1 +1 0 0 32.27±1.11 4 +1 +1 0 0 28.77±2.01 5 0 0 −1 −1 30.79±1.28 6 0 0 +1 −1 29.41±1.88 7 0 0 −1 +1 28.00±0.30 8 0 0 +1 +1 30.21±1.09 9 −1 0 0 −1 31.72±2.12 10 +1 0 0 −1 29.37±1.63 11 −1 0 0 +1 30.20±2.00 12 +1 0 0 +1 29.11±2.51 13 0 −1 −1 0 29.29±4.29 14 0 +1 −1 0 30.00±0.68 15 0 −1 +1 0 30.20±1.45 16 0 +1 +1 0 30.11±4.80 17 −1 0 −1 0 30.00±2.97 18 +1 0 −1 0 30.17±4.12 19 −1 0 +1 0 32.48±0.21 20 +1 0 +1 0 28.73±4.01 21 0 −1 0 −1 30.09±4.38 表 3 二次回归方程方差分析Table 3. ANOVA on quadratic regression equation来源
Source平方和
SS自由度
DS均方
MSF P 显著性
Significance模型
Model0.7362 14 0.0526 98.95 <0.0001 ** A 0.0878 1 0.0878 165.28 <0.0001 ** B 0.0127 1 0.0127 23.85 0.0002 ** C 0.0074 1 0.0074 13.96 0.0022 ** D 0.0248 1 0.0248 0.31 <0.0001 ** AB 0.0358 1 0.0358 0.02 <0.0001 ** AC 0.0371 01 0.0371 6.12 <0.0001 * AD 0.0044 1 0.0044 0.32 0.0118 * BC 0.0015 1 0.0015 0.08 0.1114 BD 0.0011 1 0.0011 0.02 0.1702 CD 0.0327 1 0.0327 0.03 <0.0001 ** A2 0.1299 1 0.1299 169.26 <0.0001 ** B2 0.1974 1 0.1974 9.19 <0.0001 ** C2 0.2045 1 0.2045 127.92 <0.0001 ** D2 0.2302 1 0.2302 37.54 <0.0001 ** 残差
Residual0.0074 14 0.0005 失拟项
Lack of fit0.0047 10 0.0005 0.69 0.7126 纯误差
Pure error0.0027 4 0.0007 总和
Cor total0.7436 28 R2=0.9900 R2adj=0.9800 *表示差异显著(P<0.05),**表示差异极显著(P<0.01)。
* indicates significant difference at P<0.05; ** extremely significant at P<0.01.Y=3.34−0.086A+0.21B−0.098C+0.020D+0.0092AB−0.16AC−0.036AD+0.018BC+0.01BD−0.011CD+0.65A2+0.15 B2+0.56C2+0.31D2。 通过表3方差分析表可知,模型P<0.01,说明回归方程在0.01水平极显著,表明试验设计可靠;失拟项P>0.05,说明所得方程与实际拟合非正常误差所占比例较小,可用该模型和回归方程来预测试验数据。R2=0.9900,R2adj=0.9800,进一步说明该模型拟合度较好,可信度高,约98.00%的鹿茸菇液体菌种生物量可用该模型进行预测[19]。A、B、C、D的P值小于0.01,说明其对菌丝生物量有极显著影响。AB、CD之间交互作用对菌丝生物量有极显著影响,AC、AD有显著影响,BD、BC无显著影响。比较F值大小可知,影响菌丝生物量的因素主次顺序为A全麦粉>B花生饼粉>C磷酸氢二钾>D硫酸镁,交互项影响顺序为AC>AD>BC>CD>AB=BD。
响应曲面图可更加直观地反映两两因素之间交互作用情况,等高线的形状为椭圆形表示交互作用明显,圆形则表示交互作用不明显[20]。在全麦粉质量浓度为45~52 g·L−1与花生饼粉质量浓度为21.2~23.5 g·L−1、全麦粉质量浓度为45.0~51.5 g·L−1与K2HPO4 质量浓度为1.9~2.2 g·L−1、全麦粉质量浓度为45.2~51.5 g·L−1与MgSO4·7H2O 质量浓度为1.8~2.1 g·L−1,菌丝生物量均出现极大值(图4-A~C);全麦粉与花生饼粉、碳酸氢二钾均存在显著的交互作用(表3)。其余因素之间几乎不存在交互作用,主要由单因素自身决定菌丝生物量(表3、图4)。
2.5 响应面验证试验
鹿茸菇液体菌种发酵培养基响应面优化试验中,最佳组合为全麦粉47.84 g·L−1、花生饼粉22.39 g·L−1、K2HPO4 2.04 g·L−1、MgSO4·7H2O 1.97 g·L−1,预测菌丝生物量为33.60 g·L−1。考虑实际情况,将各组用量简化为全麦粉47 g·L−1、花生饼粉22 g·L−1、K2HPO4 2.00 g·L−1、MgSO4·7H2O 2.00 g·L−1,此条件下的菌丝生物量为(32.81±1.10) g·L−1。实际值与预测值相差较小,说明本研究建立的模型可对菌丝生物量进行预测,试验设计有效可靠。
2.6 液体菌种与固体菌种出菇试验
如表4所示,液体菌种制种周期仅需16 d,较固体菌种缩短了48 d。液体菌种接种后存在多个萌发位点,2 d后观察到菌丝萌发;固体菌种是从接种块向四周生长,萌发点较少,需4 d才能萌发。鹿茸菇菌丝在栽培基质中萌发时间长,发菌较慢,较其他食用菌更易被杂菌污染,抗杂菌能力更弱,污染率高,后期出菇产量低甚至不出菇。固体菌种萌发时间长、菌丝生长速率低,污染率达29.17%,远高于液体菌种。液体菌种在菌种制备和栽培周期方面均具有优势。
表 4 液体菌种和固体菌种栽培过程及出菇比较Table 4. Comparison of fermentation processes andmushroom fruiting using liquid and solid media指标
Index液体菌种
Liquid strain固体菌种
Solid strain原种培养时间 Original culture time/d 8 24 栽培种培养时间 Culture spawn incubation time/d 8 40 制种周期 Seed production cycle/d 16 64 菌丝萌发时间 Time of mycelium germination/d 2 4 满袋时间 Bags full time/d 42 45 污染率 Pollution rate/% 8.33 29.17 现蕾时间 Budding time/d 7 9 采收时间 Harvest time/d 25 27 单包产量 Single package output/g 500.89±20.12 432.00±25.36 子实体整齐度 Fruiting body uniformity +++ ++ 菇长 Length/cm 12.82±1.19 11.76±0.83 菇径 Size/mm 12.37±1.66 17.36±1.71 盖高 Height/mm 6.86±1.35 10.72±1.85 盖径 Diameter/mm 20.06±3.77 26.10±3.94 +++表示子实体整齐度较一致; ++表示子实体整齐度一致。
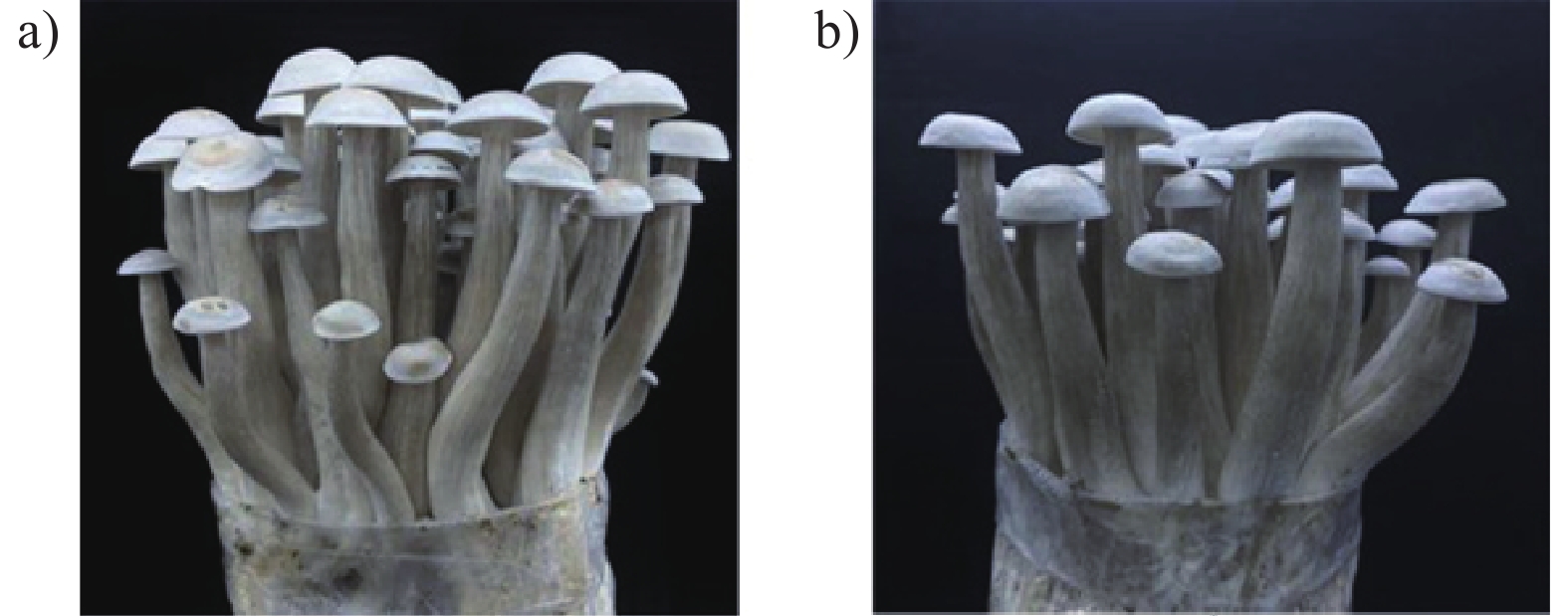
+++shows uniform fruiting body formation; ++acceptable fruiting body uniformity.如表4与图5所示,接种液体菌种的子实体整齐度较一致,单包产量和成品菇数量更高,产量较固体菌种提高15.74%。子实体农艺性状方面,接种液体菌种的子实体菇长略高于固体,而固体菌种的菇径、盖高和盖径均高于液体菌种,菌柄粗、菌盖大会影响荷叶离褶伞的口感,降低商品价值。
![]() 图 5 子实体的外观形态a):接种液体菌种的菌包;b):接种固体菌种的菌包。Figure 5. Appearance of mushroom fruiting bodiesa): Mushroom packet for inoculation of mycelia cultured in liquid medium; b): mushroom packet for inoculation of mycelia cultured in solid medium.
图 5 子实体的外观形态a):接种液体菌种的菌包;b):接种固体菌种的菌包。Figure 5. Appearance of mushroom fruiting bodiesa): Mushroom packet for inoculation of mycelia cultured in liquid medium; b): mushroom packet for inoculation of mycelia cultured in solid medium.综合分析液体菌种和固体菌种的培养周期、出菇产量、子实体农艺性状等方面,液体菌种表现优异。因此采用本研究的培养基配方所制备的液体菌种可用于鹿茸菇工厂化栽培。
3. 讨论与结论
本研究经过分析4种廉价的复合碳源与2种昂贵的单一碳源、7种廉价的复合氮源与1种昂贵的复合氮源,发现廉价型的全麦粉与花生饼粉具有作为鹿茸菇液体菌种发酵主料的潜质。此外,本项研究发现鹿茸菇菌丝可利用的碳源、氮源种类极广,但更倾向于利用复合型碳氮源,与魏生龙等[14]研究结果较一致 。在本研究中,玉米粉与小麦粉作为碳源时,鹿茸菇菌丝生物量无显著性差异,该项结果与席亚丽等[18]的研究结果较一致 ,但本研究结果表明全麦粉作为碳源培养鹿茸菇液体菌丝的效果是小麦粉的2.2倍,说明麦壳和麸皮也具有促进液体菌丝生长发育的功能[17]。分析无机盐对鹿茸菇菌丝生长的影响,发现除Co2+与Cu2+显著抑制菌球生长之外,其他供试离子均具有正效应。该研究结果与魏生龙等[14]的研究结果存在一定差异,原因可能在于鹿茸菇平板菌丝与液体菌丝生长发育对离子的吸收利用情况不同[13]。在栽培过程中,鹿茸菇菌丝在栽培料中的快速定植是极为重要的一个环节,菌丝定植时间过长,容易出现高污染,这与程继红[21]所描述的情况极为相似。本研究利用单因素-响应面优化的培养基配方,在摇瓶培养时收获菌丝生物量可达32.81±1.10 g·L−1,该值是张凌珊[16]研究结果的40余倍,应用该配方制备的液体菌种与传统固体菌种相比,制种周期、污染率均大幅下降,单包产量提高15.74%,具有较大的生产应用潜能。
本研究结果最优配方为全麦粉47 g·L−1、花生饼粉22 g·L−1、KH2PO4 2.00 g·L−1、MgSO4·7H2O 2.00 g·L−1,当发酵培养至第8 d时,菌丝生物量达到最大值(32.81±1.10g·L−1),该配方可获得高质量的生产用种。尽管如此,该液体菌种配方优化尚处于发酵优化的小试阶段,后续关于接种量、发酵罐培养方式等技术参数有待进一步优化。此外,本研究结果表明以液体发酵方式制备鹿茸菇生产用种适合鹿茸菇工厂化生产。
-
![]()
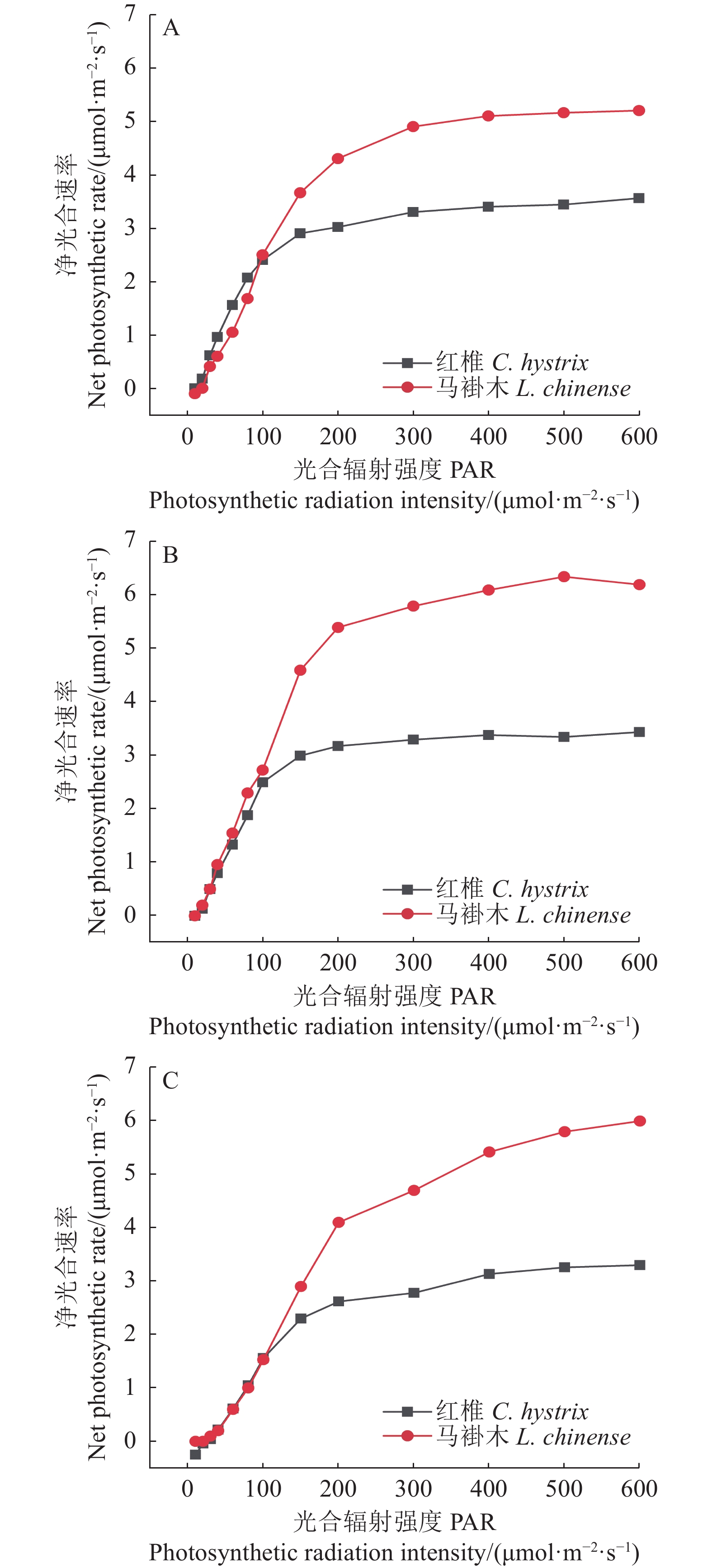
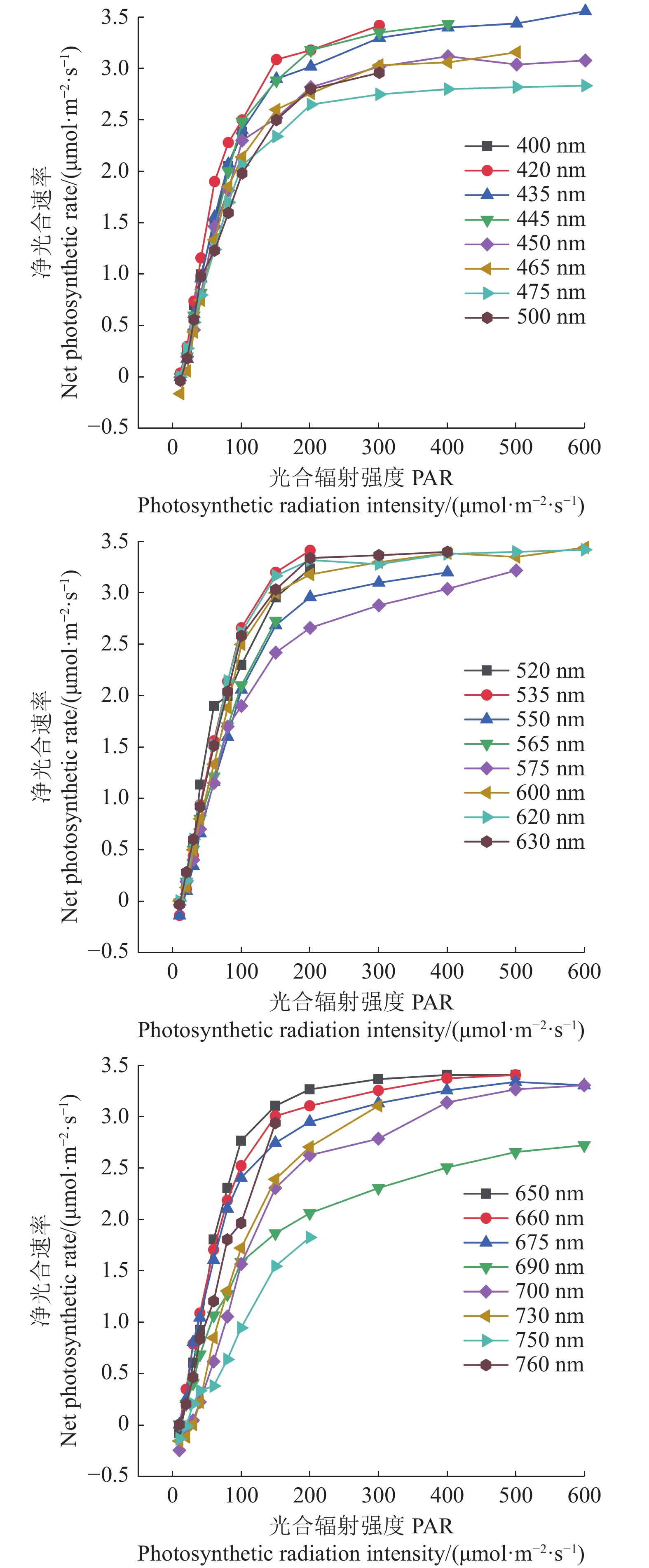
图 3 红椎和马褂木对3种不同波长的光合响应曲线
A、B、C分别表示波长为435、600、700 nm。
Figure 3. LRCs of C. hystrix and L. chinense exposed to lights of 3 different wavelengths
A, B and C represent wavelengths of 435 nm, 600 nm and 700 nm respectively.
![]()
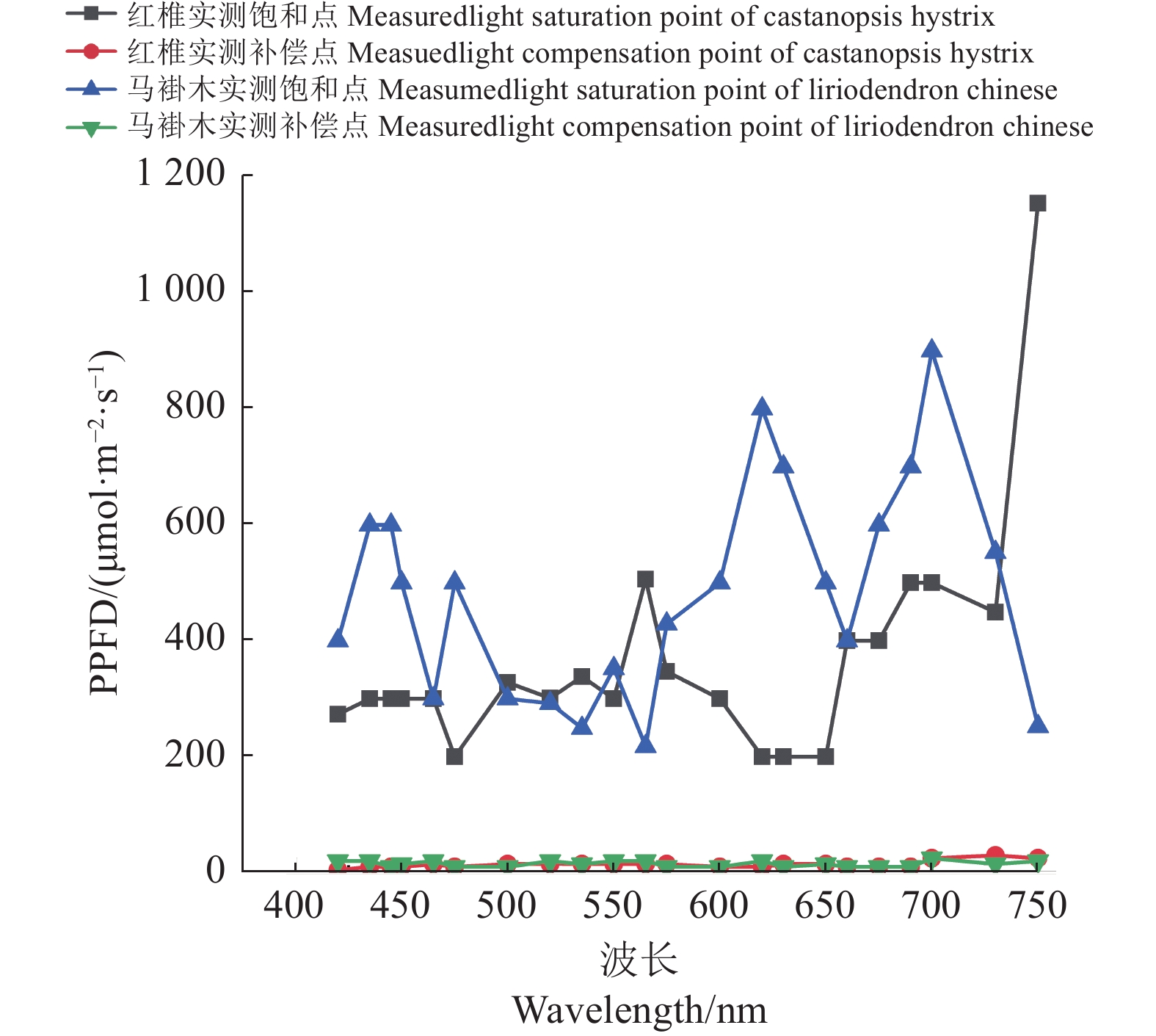
图 4 红椎与马褂木的饱和点曲线和补偿点曲线
Figure 4. Light saturation and compensation curves of C. hystrix and L. chinense
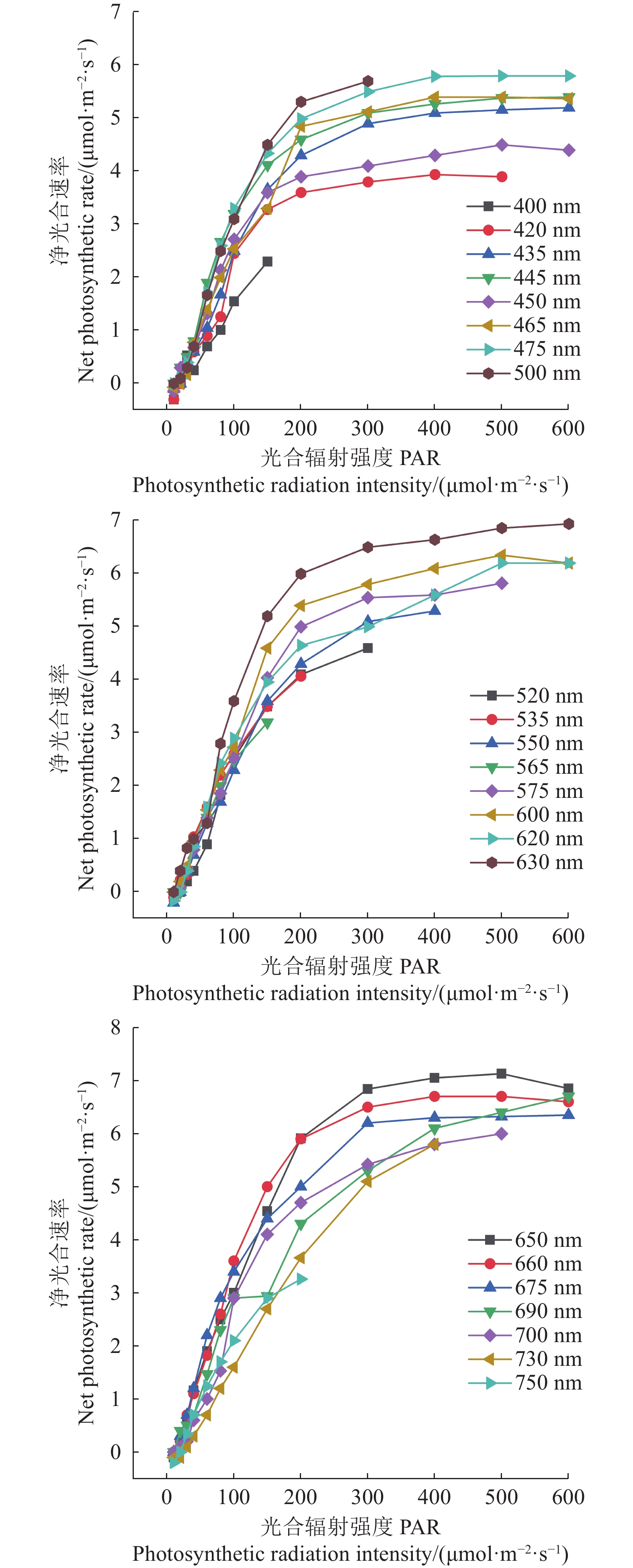
表 1 多种单色光发生器产生的22种单色光
Table 1 Twenty-two monochromatic lights produced by light generating device
波长
Wavelength λ/nm半峰全宽
Full width of
half peak ∆λ/nm波长
Wavelength λ/nm半峰全宽
Full width of
half peak ∆λ/nm420 13.9 575 22.6 435 17.9 600 20.3 445 12.9 620 17.9 450 19.4 630 17 465 15.6 650 20.5 475 25 660 11.8 500 24.2 675 26.3 520 20.5 690 26.6 535 18.7 700 33.7 550 22.4 730 28.2 565 15.7 750 28.8  下载: 导出CSV
下载: 导出CSV
表 2 红椎的光合响应曲线对应22个单色光的参数值
Table 2 Measurements related to 22 monochromatic lights on LRC of C. hystrix
波长
Wavelength
λ/nmPnb Ic/
(μmol·m−2·s−1)Ic′/
(μmol·m−2·s−1)αλ/
(m2·s−1·μmol−1)R2 波长
Wavelength
λ/nmPnb Ic/
(μmol·m−2·s−1)Ic′/
(μmol·m−2·s−1)αλ/
(m2·s−1·μmol−1)R2 420 3.45 5 12 0.015 0.995 2 575 3.14 15 14 0.01 0.995 8 435 3.47 10 13 0.013 0.996 4 600 3.45 10 15 0.013 0.990 9 445 3.5 10 14 0.012 0.993 8 620 3.45 10 14 0.015 0.990 1 450 3.09 10 14 0.013 0.993 6 630 3.51 15 14 0.013 0.993 465 3.12 15 17 0.013 0.996 8 650 3.44 15 15 0.016 0.991 7 475 2.83 10 12 0.013 0.997 2 660 3.37 10 12 0.015 0.998 500 3.13 15 12 0.011 0.995 8 675 3.28 10 11 0.014 0.996 3 520 3.51 15 13 0.013 0.985 4 690 2.81 10 6 0.007 4 0.988 7 535 3.95 15 16 0.012 0.993 1 700 3.33 25 25 0.007 8 0.991 5 550 3.31 15 17 0.011 0.994 7 730 3.59 30 24 0.007 8 0.987 4 565 4.47 15 12 0.006 9 0.998 750 5.64 25 20 0.002 2 0.987 5
下载: 导出CSV
表 3 红椎的准饱和点光强实测值
Isat 与理论计算值I′sat 的比较(p′n=0.001 )Table 3 Measured
Isat and theoreticalI′sat at quasi-saturation point of C. hystrix(p′n=0.001 )波长
Wavelength λ/nmIsat/
(μmol·m−2·s−1)Isat’/
(μmol·m−2·s−1)波长
Wavelength λ/nmIsat/
(μmol·m−2·s−1)Isat’/
(μmol·m−2·s−1)420 —— 273 575 —— 347 435 300 311 600 300 313 445 300 316 620 200 284 450 300 293 630 200 301 465 300 299 650 200 264 475 200 287 660 400 276 500 —— 328 675 400 287 520 —— 301 690 500 416 535 —— 338 700 500 441 550 300 342 730 —— 449 565 —— 506 750 —— 1154
下载: 导出CSV
表 4 马褂木的补偿响应系数、修正系数、补偿比率系数和补偿点光强
Table 4 Compensation response coefficient, correction coefficient, compensation ratio coefficient, and light intensity at compensation point of L. chinense
波长
Wavelength
λ/nmηλ
(10−3)βλ
(10−3)γλ
(10−3)Ic/
(μmol·m−2·s−1)Ic’/
(μmol·m−2·s−1)R2 波长
Wavelength
λ/nmηλ
(10−3)βλ
(10−3)γλ
(10−3)Ic/
(μmol·m−2·s−1)Ic’/
(μmol·m−2·s−1)R2 420 37 1.3 1.8 20 21 0.982 7 575 42 0.86 2 10 17 0.991 1 435 44 0.52 3.9 20 20 0.991 6 600 52 0.62 3.3 10 16 0.990 4 445 61 0.48 6.2 10 15 0.992 7 620 57 0.2 6.2 20 18 0.996 2 450 56 0.43 7.5 15 17 0.988 630 66 0.38 5.3 10 16 0.984 5 465 39 1.1 1.1 20 18 0.992 9 650 55 0.61 3.1 15 16 0.994 7 475 62 0.47 5.6 10 18 0.991 6 660 57 0.9 2.6 10 15 0.995 3 500 46 1.8 9.1×10−8 10 18 0.994 9 675 67 0.31 6.5 10 14 0.996 5 520 36 1.8 8.4×10−7 20 21 0.988 8 690 36 0.35 2.5 10 11 0.994 4 535 41 2 0.78 15 14 0.995 700 36 0.33 2.9 25 33 0.993 550 34 1.4 0.28 20 19 0.998 3 730 25 0.92 1.7×10−7 15 25 0.996 8 565 38 2.4 6.3×10−7 20 16 0.991 8 750 35 1.8 0.99 20 18 0.999 1
下载: 导出CSV
表 5 马褂木饱和点光强的实测值与理论计算值比较(
p′n=0 )Table 5 Measured and theoretical values of light intensity at saturation point of L. chinense (
p′n=0 )波长
Wavelength λ/nmIsat/
(μmol·m−2·s−1)Isat’/
(μmol·m−2·s−1)波长
Wavelength λ/nmIsat/
(μmol·m−2·s−1)Isat’/
(μmol·m−2·s−1)420 400 324 575 —— 429 435 600 521 600 500 478 445 600 465 620 800 808 450 500 472 630 700 573 465 300 385 650 500 494 475 500 491 660 400 390 500 300 281 675 600 594 520 —— 292 690 700 755 535 —— 249 700 900 780 550 —— 352 730 —— 553 565 —— 218 750 —— 252
下载: 导出CSV
-
[1] 刘文科, 查凌雁. 植物工厂植物光质生理及其调控[M]. 北京: 中国农业科学技术出版社, 2019: 2-10. [2] 刘银春, 吴金华, 吴运铨. 一种植物特征光谱实验箱及其实验方法ZL201310230571. X[P]. 2015-05-06. [3] 杨其长. 植物工厂[M]. 北京: 清华大学出版社, 2019: 3-20. [4] BIAN Z H, CHENG R F, YANG Q C, et al. Continuous light from red, blue, and green light-emitting diodes reduces nitrate content and enhances phytochemical concentrations and antioxidant capacity in lettuce [J]. Journal of the American Society for Horticultural Science, 2016, 141(2): 186−195. DOI: 10.21273/JASHS.141.2.186
[5] 樊晚林, 温振英, 周雨琦, 等. 3种野牡丹属植物的光合特性 [J]. 森林与环境学报, 2019, 39(2):188−193. FAN W L, WEN Z Y, ZHOU Y Q, et al. Photosynthetic characteristics of 3 Melastoma L. species [J]. Journal of Forest and Environment, 2019, 39(2): 188−193.(in Chinese)
[6] 张涛, 王瑞敏, 陈燕琼, 等. 8种绿化树种对城市夜间照明的生理适应性 [J]. 森林与环境学报, 2019, 39(4):424−430. ZHANG T, WANG R M, CHEN Y Q, et al. Physiological adaptations of eight greening-trees in an urban area under night lighting [J]. Journal of Forest and Environment, 2019, 39(4): 424−430.(in Chinese)
[7] BIAN Z H, CHENG R F, WANG Y, et al. Effect of green light on nitrate reduction and edible quality of hydroponically grown lettuce (Lactuca sativa L. ) under short-term continuous light from red and blue light-emitting diodes [J]. Environmental and Experimental Botany, 2018, 153: 63−71. DOI: 10.1016/j.envexpbot.2018.05.010
[8] 卢思, 罗梅秀, 刘丹, 等. 不同光质对三叶青茎叶显微结构和激素的影响 [J]. 森林与环境学报, 2019, 39(1):15−20. LU S, LUO M X, LIU D, et al. Responses of Tetrastigma hemsleyanum microstructure and phytohormone content to different light quality [J]. Journal of Forest and Environment, 2019, 39(1): 15−20.(in Chinese)
[9] 粟春青, 蒋霞, 亢亚超, 等. 金花茶幼苗对铅胁迫的生长生理响应 [J]. 森林与环境学报, 2019, 39(5):467−474. SU C Q, JIANG X, KANG Y C, et al. Growth and physiological response of Camellia petelotii seedlings to Pb stress [J]. Journal of Forest and Environment, 2019, 39(5): 467−474.(in Chinese)
[10] 刘莉娜, 张卫强, 黄芳芳, 等. 盐胁迫对银叶树幼苗光合特性与叶绿素荧光参数的影响 [J]. 森林与环境学报, 2019, 39(6):601−607. LIU L N, ZHANG W Q, HUANG F F, et al. Effects of NaCl stress on the photosynthesis and cholorophyll fluorescence of Heritiera littoralis seedlings [J]. Journal of Forest and Environment, 2019, 39(6): 601−607.(in Chinese)
[11] 赵子豪, 宋琦, 李利, 等. 南方四季杨雌雄幼苗对镉胁迫光合生理响应的差异 [J]. 森林与环境学报, 2019, 39(2):201−207. ZHAO Z H, SONG Q, LI L, et al. Difference in photosynthesis and physiological response of male and female Populus deltoides × nigra to Cd stress [J]. Journal of Forest and Environment, 2019, 39(2): 201−207.(in Chinese)
[12] SHAO L Z, FU Y M, LIU H, et al. Changes of the antioxidant capacity in Gynura bicolor DC under different light sources [J]. Scientia Horticulturae, 2015, 184: 40−45. DOI: 10.1016/j.scienta.2014.12.010
[13] 何天友, 于增金, 沈少炎, 等. 花吊丝竹对干旱胁迫的光合和生理响应 [J]. 森林与环境学报, 2020, 40(1):68−75. HE T Y, YU Z J, SHEN S Y, et al. Photosynthetic and physiological responses of Dendrocalamus minor var. amoenus to drought stress [J]. Journal of Forest and Environment, 2020, 40(1): 68−75.(in Chinese)
[14] BIAN Z H, YANG Q C, LIU W K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review [J]. Journal of the Science of Food and Agriculture, 2015, 95(5): 869−877. DOI: 10.1002/jsfa.6789
[15] 李义博, 宋贺, 周莉, 等. C4植物玉米的光合-光响应曲线模拟研究 [J]. 植物生态学报, 2017, 41(12):1289−1300. DOI: 10.17521/cjpe.2017.0158 LI Y B, SONG H, ZHOU L, et al. Modeling study on photosynthetic-light response curves of a C4 plant, maize [J]. Chinese Journal of Plant Ecology, 2017, 41(12): 1289−1300.(in Chinese) DOI: 10.17521/cjpe.2017.0158
[16] 闫小红, 尹建华, 段世华, 等. 四种水稻品种的光合光响应曲线及其模型拟合 [J]. 生态学杂志, 2013, 32(3):604−610. DOI: 10.13292/j.1000-4890.2013.0084 YAN X H, YIN J H, DUAN S H, et al. Photosynthesis light response curves of four rice varieties and model fitting [J]. Chinese Journal of Ecology, 2013, 32(3): 604−610.(in Chinese) DOI: 10.13292/j.1000-4890.2013.0084
[17] 李佳, 刘济明, 文爱华, 等. 米槁幼苗光合作用及光响应曲线模拟对干旱胁迫的响应 [J]. 生态学报, 2019, 39(3):913−922. LI J, LIU J M, WEN A H, et al. Simulated photosynthetic responses of Cinnamomum migao during drought stress evaluated using Light-response Models [J]. Acta Ecologica Sinica, 2019, 39(3): 913−922.(in Chinese)
[18] 叶子飘. 光合作用对光和CO2响应模型的研究进展 [J]. 植物生态学报, 2010, 34(6):727−740. DOI: 10.3773/j.issn.1005-264x.2010.06.012 YE Z P. A review on modeling of responses of photosynthesis to light and CO2 [J]. Chinese Journal of Plant Ecology, 2010, 34(6): 727−740.(in Chinese) DOI: 10.3773/j.issn.1005-264x.2010.06.012
[19] 叶子飘, 于强. 一个光合作用光响应新模型与传统模型的比较 [J]. 沈阳农业大学学报, 2007, 38(6):771−775. DOI: 10.3969/j.issn.1000-1700.2007.06.001 YE Z P, YU Q. Comparison of A new model of light response of photosynthesis with traditional models [J]. Journal of Shenyang Agricultural University, 2007, 38(6): 771−775.(in Chinese) DOI: 10.3969/j.issn.1000-1700.2007.06.001
[20] 秦惠珍, 韦霄, 唐健民, 等. 东兴金花茶和长尾毛蕊茶光合响应曲线拟合模型比较研究 [J]. 江苏农业科学, 2020, 48(15):165−170. DOI: 10.15889/j.issn.1002-1302.2020.15.029 QIN H Z, WEI X, TANG J M, et al. Comparative study on fitting models of photosynthetic response curves between Camellia tunghinensis Chang and Camellia caudata Wall [J]. Jiangsu Agricultural Sciences, 2020, 48(15): 165−170.(in Chinese) DOI: 10.15889/j.issn.1002-1302.2020.15.029
[21] 钱一凡, 廖咏梅, 权秋梅, 等. 4种光响应曲线模型对3种十大功劳属植物的实用性 [J]. 植物研究, 2014, 34(5):716−720. DOI: 10.7525/j.issn.1673-5102.2014.05.021 QIAN Y F, LIAO Y M, QUAN Q M, et al. Practicability of four models of light-response curves of photosynthesis to three Mahonia species [J]. Bulletin of Botanical Research, 2014, 34(5): 716−720.(in Chinese) DOI: 10.7525/j.issn.1673-5102.2014.05.021
[22] KORSAKOVA S, PLUGATAR Y, ILNITSKY O, et al. A research on models of the photosynthetic light response curves on the example of evergreen types of plants [J]. Agronomy Research, 2019, 17(2): 518−539.
[23] 刘银春, 段亚凡, 林火养, 等. 双驼峰光谱发光装置及可输出多种单色光的装置: CN209102595U[P]. 2019-07-12. -
期刊类型引用(5)
1. 周静,张馨,赵兴秀,何义国,陈雨,张靖. 长裙竹荪液体培养基优化. 食用菌. 2024(03): 9-13+16 .  百度学术
百度学术
2. 金文松,李建华,单灿灿,吴涛锐,程泳春,程利纹,李佳欢,孙淑静,胡开辉. 鹿茸菇菌包后熟指标与出菇产量间的相关性. 福建农林大学学报(自然科学版). 2024(04): 563-569 . 百度学术
3. 彭汇涵,于海玲,王明元,杨苗,万雨欣. 鹿茸菇液体菌种培养过程中生理指标的变化规律. 华侨大学学报(自然科学版). 2024(05): 667-672 . 百度学术
4. 龚娜,刘国丽,陈珣,马晓颖,肇莹,肖军. 一株野生肺形侧耳的鉴定及其液体发酵培养基的优化. 浙江农业学报. 2024(11): 2535-2545 . 百度学术
5. 陈珣,肇莹,龚娜,刘国丽,马晓颖,张敏,肖军. 基于响应面法优化滑菇液体菌种发酵培养基配方. 南方农业学报. 2024(12): 3625-3635 . 百度学术
其他类型引用(0)

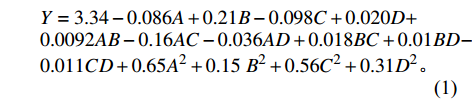

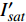
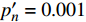
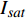
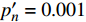
计量
- 文章访问数: 377
- HTML全文浏览量: 199
- PDF下载量: 21
- 被引次数: 5
