

Preparation of Monoclonal Antibodies Against CD25 of Porcine Tregs
-
摘要:目的
制备猪源CD25蛋白单克隆抗体,为猪源Treg细胞的检测及生物学研究提供基础。
方法将6×His猪源CD25基因片段通过同源重组的方法克隆至原核表达载体pET-28a,构建重组质粒pET28a-CD25。然后利用大肠杆菌表达系统和金属螯合亲和层析介质(Ni-NTA)预装柱对6×His-CD25重组蛋白进行表达和纯化。以纯化的重组蛋白作为抗原免疫雌性BALB/c小鼠,取免疫后小鼠的脾细胞与骨髓瘤细胞(SP2/0)进行细胞融合,经过间接ELISA筛选融合细胞并进行克隆化培养,并利用小鼠腹水法大量制备猪源CD25单克隆抗体。
结果成功获得一株由单一细胞生长且能够稳定分泌CD25单克隆抗体的杂交瘤细胞株。使用间接ELISA与Western blotting分析显示,CD25单克隆抗体不与其他蛋白产生免疫反应,抗体效价为1∶
4098000 ,表明CD25单克隆抗体特异性较强且效价高。间接免疫荧光和免疫印迹分析结果显示,CD25单克隆抗体不仅可以识别真核细胞表达的CD25蛋白,而且可以识别宿主脾脏和肺脏中的内源性CD25蛋白。结论成功制备了重组猪源CD25重组蛋白并筛选出特异性单克隆抗体,为定性或定量检测猪源Treg细胞CD25蛋白和研究猪源Treg细胞的功能奠定了基础。
Abstract:ObjectiveThe monoclonal antibody against CD25 in pigs was obtained to facilitate detecting and studying the regulatory T cells (Tregs).
MethodThe porcine 6×His tagged CD25 fragment was cloned into prokaryotic expression vector pET-28a via homologous recombination to construct the recombinant plasmid pET28a-CD25. Subsequently, the 6×His-CD25 recombinant protein was expressed in E. coli and purified on a metal chelating affinity chromatography medium (Ni-NTA) pre-packed column to be used as the antigen to immunize female BALB/c mice. Spleen cells of the immunized mice were hybridized with myeloma cells SP2/0 and screened by indirect ELISA prior to culture cloning. The target monoclonal antibody against CD25 was thus prepared using mouse ascites method.
ResultA single cell-derived hybridoma cell line with a stable expression of CD25 monoclonal antibody was successful obtained. ELISA and western blotting showed the antibody had a strong specificity with an exclusive reactivity towards the target protein, not the others, even at a high titer of 1:
4098000 . In the indirect immunofluorescence and western blotting assays, the antibody not only detected CD25 in the eukaryotic cells but also endogenous one in the host spleen and lung tissues.ConclusionThe recombinant porcine CD25 was successfully prepared and the specific monoclonal antibody obtained for qualitative and/or quantitative detection of the protein as well as further studies on the functions of porcine Tregs.
-
Keywords:
- pig /
- Treg cells /
- CD25 /
- monoclonal antibody
-
0. 引言
【研究意义】芸芥(Eruca sativa M.)系十字花科芝麻菜属植物,是重要的油料和蔬菜作物,在欧洲、非洲、中东、中亚、印度和我国甘肃与宁夏等地都有栽培[1]。土壤盐渍化及次生盐渍化是一种典型的非生物逆境胁迫因素,不仅影响作物生长发育,还对生态环境产生不良效应[2]。据不完全统计,我国将近0.2亿多hm2的农田受到盐害影响[3]。盐害可危及作物的芽期、苗期和全生育期[4],其对植物的影响主要表现为抑制种子萌发、限制幼苗生长、降低光合作用、改变新陈代谢、水分吸收失调、离子营养不平衡、氧化胁迫、产量降低等[5]。不同盐胁迫类型中,NaCl是最为常见的,但随着温室二氧化碳和工业二氧化硫排放量的增加,以及工厂废弃液的排放、各种土壤改良剂的大量施用、地壳的逐渐风化,形成越来越多的硫酸盐、碳酸盐等盐碱环境,使植物遭受的其他种类盐碱胁迫的面积比NaCl胁迫的面积更大,对植物的危害更深[6]。因此进行芸芥耐盐性评价,选育耐盐品系或品种,对利用大面积的盐碱地和保证芸芥籽供应具有重要的现实意义。【前人研究进展】芸芥与芸薹属有一定亲缘关系,具有优异抗旱、耐瘠薄、抗白粉病、耐低温等抗逆特性[7]。从已有的资料分析,在盐碱逆境中,芸芥具有一定的耐性[8],尽管有关盐胁迫对芸芥萌发、生长发育和生理特性的影响已有一些报道[9],但主要以某一种盐或某一个品种开展研究为主[10-12]。【本研究切入点】有关盐胁迫对芸芥不同品系或芸芥耐不同种类盐胁迫能力方面的报道较少。同一种植物不同品种或品系对盐碱环境的抗性存在明显差异,在萌发期种子发芽率等性状对盐、碱胁迫非常敏感[2]。【拟解决的关键问题】本试验以3个芸芥育成品系为材料,鉴定并评价不同品系在NaCl、Na2SO4、NaHCO3和Na2CO3 等4种单盐胁迫下萌发期各指标的变化,以期探明芸芥萌发期对不同种类盐碱胁迫的适应条件,为进一步的耐盐品种选育和盐碱地的改良提供一定的参考依据。
1. 材料与方法
1.1 供试材料
3个芸芥品系,包括15芸芥4、11芸芥32-1、16芸芥9,农艺性状优良,生育期120~130 d,由河西学院农业与生态工程学院农学教研室提供。试验前挑选饱满一致的种子,在70%乙醇中浸泡30 s,用40%次氯酸钠溶液消毒15 min,无菌水清洗3次,晾干后备用。
1.2 试验方法
依照国际种子检验规程,发芽采用滤纸床法,将双层滤纸用盐溶液充分饱和后置于内径100 mm的培养皿底部,之后放置种子100粒,加入不同浓度的NaCl、NaHCO3、Na2CO3和Na2SO4等4种单盐溶液进行胁迫处理。其中NaCl采用浓度为30、60、90、120、150、180、210、240、300 mmol·L−1,Na2SO4采用浓度为30、60、90、120、150 mmol·L−1,NaHCO3采用浓度为15、30、45、60 mmol·L−1,Na2CO3采用浓度5、10、15、20、25、30 mmol·L−1,分别以不加单盐溶液为对照,每个处理重复3次,置于25 ℃人工气候培养箱中培养,相对湿度为65%,光照时间与非光照时间设定均为12 h。试验过程中每天采用称重补水法按时补充水分,以保持处理期间盐浓度相对稳定。
1.3 指标测定与数据处理
发芽期间,每天记录种子发芽情况及发芽数,胚根突破种皮2 mm为发芽标准,共计7 d,连续2 天种子发芽数不变即调查结束。发芽试验进行至第10 天时,每个培养皿中随机取10个幼苗,用刻度0.5 mm的直尺从胚轴与胚根的分界处分别测定胚轴和胚根绝对长度;然后用滤纸吸干幼苗表面的水分后立即放入精确度为0.0001 g的天平上称重,计算单个幼苗的平均重量,其中:发芽势=第3天发芽的种子数量/种子总数×100%,发芽率=第7天发芽的种子数量/种子总数×100%,发芽指数=∑Gt/Dt(Gt:当天的发芽数;Dt:发芽日数)。
参考李宏等[13]、裴毅等[14]和白小明等[15]的方法:先对发芽率和盐碱胁迫强度进行相关分析,若两者存在显著相关性,则对NaCl、Na2SO4、NaHCO3和Na2CO3等4种单盐胁迫下3个芸芥品系的发芽率、发芽势、发芽指数、胚轴长、胚根长和幼苗鲜重分别进行回归统计分析。以盐浓度为自变量建立各指标的回归方程,以各指标的测定值下降到对照的50%和0时分别求得3个芸芥品系在相应指标下的耐盐临界值和耐盐极限值,最后求出同一指标下3个品系的平均耐盐临界值和耐盐极限值,以及同一品系6个指标的平均耐盐临界值和耐盐极限值,即综合评价值,并排序以判断耐盐能力。数据采用Excel 2016软件统计,采用SPSS 22.0软件中单因素方差分析的LSD法进行不同浓度处理下的差异显著性分析。
2. 结果与分析
2.1 NaCl胁迫对芸芥品系萌发期的影响
试验结果表明,在NaCl胁迫下,15芸芥4耐盐性最强,11芸芥32-1居中,16芸芥9最弱(图1)。其中图1-A表明,当NaCl为90 mmol·L−1时,16芸芥9发芽势下降为83.3%,与对照间差异达显著水平;当NaCl为150 mmol·L−1时,15芸芥4发芽势较对照显著下降,而11芸芥32-1和16芸芥9发芽势≤20%,可见,发芽初期15芸芥4品系发芽力较强;当NaCl浓度分别为300、270 mmol·L−1时,3个品系发芽势均为0。由图1-B可知,当NaCl浓度为150 mmol·L−1时,11芸芥32-1和16芸芥9发芽指数≤7,15芸芥4发芽指数为15.3,比对照低60.69%;NaCl浓度为270 mmol·L−1时,3个品系发芽指数≤3,16芸芥9的最低,仅0.4,这表明15芸芥4品系种子生命力较强。从图1-C来看,在NaCl浓度为150~300 mmol·L−1时,15芸芥4发芽率最高,11芸芥32-1次之,16芸芥9最低。由图1-D可知,当NaCl浓度为120 mmol·L−1时,15芸芥4品系幼苗鲜重为0.0289 g,与对照间达显著差异水平,而11芸芥32-1和16芸芥9幼苗鲜重与对照间无显著差异;当NaCl浓度为300 mmol·L−1时,3个品系幼苗鲜重均低于0.0001 g。由图1-E可知,当NaCl浓度为30~60 mmol·L−1时,11芸芥32-1胚轴长2.94~3.16 cm,比对照高了11.36%~19.69%;当NaCl浓度为150 mmol·L−1时,15芸芥4胚轴最长,而11芸芥32-1胚轴最短。由图1-F可知,当NaCl浓度为90~120 mmol·L−1时,15芸芥4胚根最长;当NaCl浓度为180~240 mmol·L−1时,15芸芥4胚根长较对照显著下降,11芸芥32-1胚根长为0.69~0.71 cm,16芸芥9胚根长最短。15芸芥4、11芸芥32-1和16芸芥9胚根和胚轴生长受到完全抑制时,NaCl浓度分别为300、300和270 mmol·L−1。
![]() 图 1 NaCl溶液胁迫对3个芸芥品系种子萌发性状的影响注:A.发芽势;B.发芽指数;C.发芽率;D. 幼苗鲜重;E.胚根长;F.胚轴长; 小写字母表示同一种盐不同浓度处理间5%水平差异显著(P<0.05),图2~4同。Figure 1. Effects of NaCl on germination of 3 E. sativa linesNotes:A: germination potential; B: germination index; C: germination rate; D: fresh weight; E: radical length; F: hypocotyl length. Data with different lowercase letters indicated significant difference (P<0.05). Same for Fig.2–4.
图 1 NaCl溶液胁迫对3个芸芥品系种子萌发性状的影响注:A.发芽势;B.发芽指数;C.发芽率;D. 幼苗鲜重;E.胚根长;F.胚轴长; 小写字母表示同一种盐不同浓度处理间5%水平差异显著(P<0.05),图2~4同。Figure 1. Effects of NaCl on germination of 3 E. sativa linesNotes:A: germination potential; B: germination index; C: germination rate; D: fresh weight; E: radical length; F: hypocotyl length. Data with different lowercase letters indicated significant difference (P<0.05). Same for Fig.2–4.2.2 Na2SO4胁迫对芸芥品系萌发期的影响
图2-A和2-B表明,在Na2SO4浓度90 mmol·L−1处理下,发芽势和发芽指数均显著下降,16芸芥9发芽势和发芽指数均最低,分别为3.3%和2.8,表明种子活力非常低;Na2SO4浓度为180 mmol·L−1时,发芽势和发芽指数均为0,表明参试芸芥种子发生裂变,完全失去发芽力。从图2-C可知,当Na2SO4浓度为30 mmol·L−1时,15芸芥4幼苗鲜重较对照显著下降,而11芸芥32-1幼苗鲜重较对照高了0.0056 g,表明幼苗健壮程度有所提高;Na2SO4浓度为90 mmol·L−1时,幼苗鲜重显著下降,与高、低浓度处理间均存在显著差异。
![]() 图 2 Na2SO4溶液胁迫对3个芸芥品系种子萌发性状的影响Figure 2. Effects of Na2SO4 on germination of 3 E. sativa lines
图 2 Na2SO4溶液胁迫对3个芸芥品系种子萌发性状的影响Figure 2. Effects of Na2SO4 on germination of 3 E. sativa lines由图2-A和2-C可知,30 mmol·L−1 Na2SO4处理下,发芽势和发芽率变化基本一致,11芸芥32-1测定值与对照一致,其余3个品系比对照低,且无显著差异,表明种子活力较高;Na2SO4浓度为60 mmol·L−1时,11芸芥32-1和16芸芥9发芽率均为90%,但前者发芽势较高,表明种子生命力强。发芽率为0时,15芸芥4、16芸芥9和11芸芥32-1所对应的Na2SO4浓度分别为180、120和150 mmol·L−1;当Na2SO4浓度为90 mmol·L−1时,15芸芥4、16芸芥9和11芸芥32-1等3个品系的发芽率分别为73.3%、33.3%和50.0%。
由图2-D、2-E和2-F可知,用30 mmol·L−1 Na2SO4处理,11芸芥32-1胚根长大于对照,15芸芥4和11芸芥32-1胚轴长大于对照,但无显著性差异,这表明低浓度Na2SO4盐处理,能促进15芸芥4胚轴的生长,也能促进11芸芥32-1胚轴和胚根的生长;当Na2SO4浓度为90 mmol·L−1时,3个品系胚轴长为0.38~0.83 cm,胚根长0.78~1.30 cm,16芸芥9胚轴和胚根最短,11芸芥32-1次之,15芸芥4最大,说明对16芸芥9幼苗生长产生的抑制作用最强。在150 mmol·L−1≤Na2SO4浓度≤180 mmol·L−1处理下,胚根长、胚轴长和幼苗鲜重均为0,说明幼苗形态建成受到完全限制。
综上,在Na2SO4胁迫下,15芸芥4耐盐性最强,11芸芥32-1居中,16芸芥9最弱。
2.3 NaHCO3胁迫对芸芥品系萌发期的影响
由图3-A、3-B和3-C可知,在15 mmol·L−1 NaHCO3处理下,11芸芥32-1发芽势比对照高了3.41%,发芽率与对照一致,发芽指数略低于对照,这表明低浓度NaHCO3处理能提高11芸芥32-1种子的发芽速度,种子活力变化小,其余2个品系发芽势、发芽率均降低,但都在90%以上;NaHCO3浓度为30 mmol·L−1时,发芽率较对照显著下降,11芸芥32-1发芽率下降幅度最大,表明其种子生活力受到较强的抑制效应。在60 mmol·L−1 NaHCO3处理下,除15芸芥4外,其余2个品系发芽势、发芽率和发芽指数均为0,表明种子完全失去生命力。发芽率为0时,15芸芥4、11芸芥32-1和16芸芥9所对应的NaHCO3浓度分别为75、60、60 mmol·L−1;当NaHCO3浓度为60 mmol·L−1时,15芸芥4种子仍有萌发能力,而16芸芥9和11芸芥32-1发芽率均为0。
![]() 图 3 NaHCO3溶液胁迫对3个芸芥品系种子萌发性状的影响Figure 3. Effects of NaHCO3 on germination of 3 E. sativa lines
图 3 NaHCO3溶液胁迫对3个芸芥品系种子萌发性状的影响Figure 3. Effects of NaHCO3 on germination of 3 E. sativa lines由图3-D、3-E和3-F可知,NaHCO3浓度为15 mmol·L−1时,11芸芥32-1幼苗鲜重和胚轴长分别较对照高了14.16%和7.43%,其余2个品系的均比对照值下降,3个品系胚根长均显著下降;NaHCO3浓度为45 mmol·L−1时,11芸芥32-1幼苗鲜重、胚轴长和胚根长均为0,15芸芥4次之,而16芸芥9三个指标均最高;在75 mmol·L−1 NaHCO3浓度处理下,3个品系幼苗鲜重、胚轴长和胚根长均最低,表明高浓度NaHCO3胁迫不能促成幼苗形态的建成。
综上,在NaHCO3处理下,16芸芥9耐盐性最强,15芸芥4居中,11芸芥32-1最弱。
2.4 Na2CO3胁迫对芸芥品系萌发期的影响
由图4-A、4-B和4-C可知,当Na2CO3浓度为10 mmol·L−1时,3个品系发芽率均在86.7%以上,当Na2CO3浓度为20 mmol·L−1时,发芽率显著下降,11芸芥32-1、16芸芥9和15芸芥4分别较对照下降了90%、81.4%和80%,除15芸芥4之外,其余品系发芽势为0,11芸芥32-1发芽指数最低,仅0.38,这说明15芸芥4的种子活力较强。另外,当Na2CO3浓度分别为35、30、25 mmol·L−1时,15芸芥4、16芸芥9和11芸芥32-1的发芽率均为0;当Na2CO3浓度为25 mmol·L−1时,15芸芥4和16芸芥9种子仍有萌发能力,发芽率分别为17%和6.7%,而11芸芥32-1发芽率为0,出现这种结果,可能是11芸芥32-1贮藏时间较长、抗逆性下降,也可能是该品系耐Na2CO3胁迫的能力较差;当Na2CO3浓度为30 mmol·L−1时,仅15芸芥4能发芽,发芽率低至7%。
![]() 图 4 Na2CO3溶液胁迫对3个芸芥品系种子萌发性状的影响Figure 4. Effects of Na2CO3 on germination of 3 E. sativa lines
图 4 Na2CO3溶液胁迫对3个芸芥品系种子萌发性状的影响Figure 4. Effects of Na2CO3 on germination of 3 E. sativa lines由图4-D、4-E和4-F可知,当Na2CO3浓度为5 mmol·L−1时,15芸芥4的幼苗鲜重、胚根长、胚轴长分别较对照升高了27%、21.99%和0.9%;当Na2CO3浓度为10~15 mmol·L−1时,参试品系幼苗鲜重、胚根长和胚轴长均较对照显著下降,胚根长下降幅度最大;20 mmol·L−1 Na2CO3胁迫完全抑制了11芸芥32-1胚根,甚至不能形成幼苗。30 mmol·L−1 Na2CO3胁迫抑制了参试品系幼苗形态建成。
综上,在Na2CO3胁迫下,15芸芥4耐盐性最强,16芸芥9居中,11芸芥32-1最弱。
2.5 4种钠盐胁迫下3个芸芥品系萌发期耐盐临界值和极限值差异
由表1可知,NaCl胁迫下参试品系6个指标耐盐临界值为128~185 mmol·L−1,平均值158 mmol·L−1;15芸芥4耐NaCl界值为141~244 mmol·L−1,11芸芥32-1的耐NaCl临界值为101~223 mmol·L−1,16芸芥9不同指标耐NaCl临界值为68~178 mmol·L−1;3个品系平均耐NaCl临界值和耐盐极限值以发芽率最大(207、283 mmol·L−1),幼苗鲜重位居第二(189、275 mmol·L−1),胚根长最小(112、195 mmol·L−1);3个品系中,6个指标平均NaCl盐临界值和耐NaCl极限值均以15芸芥4最大,11芸芥32-1第二,16芸芥9最小,表明3个芸芥品系耐NaCl胁迫强弱依次为:15芸芥4>11芸芥32-1>16芸芥9,15芸芥4耐NaCl适应性较广。
表 1 NaCl胁迫下3个芸芥品系种子萌发期耐盐临界值和极限值Table 1. Tolerance thresholds and limits of 3 E. sativa lines on NaCl品系
Lines耐盐临界值Salt tolerance threshold/(mmol·L−1) 耐盐极限值Salt tolerance limit/(mmol·L−1) 品系排序
Line sequenceA B C D E F 平均值
MeanA B C D E F 平均值
Mean15芸芥4 15yunjie4 244 173 141 212 174 168 185 336 239 209 306 247 245 264 1 11芸芥32-1 11 yunjie32-1 199 143 132 223 170 101 161 271 200 198 327 240 181 236 2 16芸芥9 16yunjie9 178 127 122 133 141 68 128 243 185 188 191 218 160 198 3 平均值 Mean 207 148 132 189 162 112 158 283 208 198 275 235 195 233 指标排序 Index sequence 1 4 5 2 3 6 1 4 5 2 3 6 注:A.发芽势;B.发芽指数;C.发芽率;D. 幼苗鲜重;E.胚根长;F.胚轴长. 表2~4同.
Notes: A: germxsination potential; B: germination index; C: germination rate; D: fresh weight; E: radical length; F: hypocotyl length. Same for Table 2–4.由表2可知,Na2SO4胁迫下,3个芸芥品系平均耐Na2SO4临界值和极限值以发芽率最大,分别为107和169 mmol·L−1,胚轴长第二,分别为86和120 mmol·L−1,胚根长最小,分别为65和102 mmol·L−1;15芸芥4耐Na2SO4临界值为59~120 mmol·L−1,11芸芥32耐Na2SO4临界值为72~104 mmol·L−1,16芸芥9耐Na2SO4临界值为61~97 mmol·L−1;3个品系中,6个指标的平均耐Na2SO4临界值和极限值均以15芸芥4最大,11芸芥32-1第二,16芸芥9最小,这表明3个芸芥品系耐Na2SO4胁迫依次为:15芸芥4>11芸芥32-1>16芸芥9,15芸芥4耐Na2SO4适应性较广。
表 2 Na2SO4胁迫下3个芸芥品系种子萌发期耐盐临界值和极限值Table 2. Tolerance thresholds and limits of 3 E. sativa lines on Na2SO4品系
Lines耐盐临界值Salt tolerance threshold/(mmol·L−1) 耐盐极限值Salt tolerance limit/(mmol·L−1) 品系排序
Line sequenceA B C D E F 平均值
MeanA B C D E F 平均值
Mean15芸芥4 15yunjie4 120 84 81 84 99 59 88 178 121 120 121 135 105 130 1 11芸芥32-1 11 yunjie32-1 104 82 72 88 84 74 84 168 118 112 125 125 112 127 2 16芸芥9 16yunjie9 97 65 60 81 74 61 73 161 91 87 114 102 90 108 3 平均值 Mean 107 77 71 84 86 65 82 169 110 106 120 120 102 121 指标排序 Index sequence 1 4 5 3 2 6 1 3 4 2 2 5 由表3可知,NaHCO3胁迫下,3个品系耐NaHCO3临界值为65~107 mmol·L−1,平均值34 mmol·L−1,6个指标的耐NaHCO3极限值为36~67 mmol·L−1,平均值52 mmol·L−1;15芸芥4耐NaHCO3临界值为21~39 mmol·L−1,11芸芥32-1耐NaHCO3临界值为17~34 mmol·L−1,16芸芥9耐NaHCO3临界值为23~72 mmol·L−1;3个品系平均耐NaHCO3临界值和极限值以幼苗鲜重最大,分别是46、67 mmol·L−1,胚根长最小,分别是20、36 mmol·L−1;3个品系中,6个指标平均耐NaHCO3临界值和极限值以16芸芥9最大,15芸芥4第二,11芸芥32-1最小,表明3个芸芥品系耐NaHCO3胁迫依次为:16芸芥9>15芸芥4>11芸芥32-1,16芸芥9耐NaHCO3适应性较广。
表 3 NaHCO3胁迫下3个芸芥品系种子萌发期耐盐临界值和极限值Table 3. Tolerance thresholds and limits of 3 E. sativa lines on NaHCO3品系
Lines耐盐临界值Salt tolerance threshold/(mmol·L−1) 耐盐极限值Salt tolerance limit/(mmol·L−1) 品系排序
Line sequenceA B C D E F 平均值
MeanA B C D E F 平均值
Mean15芸芥4 15yunjie4 36 34 27 39 36 21 32 65 50 45 56 53 41 52 2 11芸芥32-1 11 yunjie32-1 34 25 25 28 28 17 26 62 35 35 37 38 29 39 3 16芸芥9 16yunjie9 40 27 28 72 64 23 42 67 38 40 107 95 37 64 1 平均值 Mean 37 29 27 46 43 20 34 65 41 40 67 62 36 52 指标排序 Index sequence 3 4 5 1 2 6 2 4 5 1 3 6 由表4可知,Na2CO3胁迫下,3个品系耐Na2CO3临界值和极限值的平均值分别为14.8、21 mmol·L−1,变化范围分别为12.3~16.7 mmol·L−1、18~23.3 mmol·L−1;15芸芥4耐Na2CO3临界值为13~21 mmol·L−1,11芸芥32耐Na2CO3临界值为10~16 mmol·L−1,16芸芥9耐Na2CO3临界值为12~19 mmol·L−1;3个品系平均耐Na2CO3临界值和极限值以胚轴长的最大,分别为16.7、23.3 mmol·L−1,胚根长最小,分别为12.3、18 mmol·L−1;3个品系中,平均耐Na2CO3临界值和极限值均以15芸芥4最大,16芸芥9第二,11芸芥32-1最小,这表明3个芸芥品系耐Na2CO3胁迫依次为:15芸芥4>16芸芥9>11芸芥32-1,15芸芥4耐Na2CO3适应性较广。
表 4 Na2CO3胁迫下3个芸芥品系种子萌发期耐盐临界值和极限值Table 4. Tolerance thresholds and limits of 3 E. sativa lines on Na2CO3品系
Lines耐盐临界值Salt tolerance threshold/(mmol·L−1) 耐盐极限值Salt tolerance limit/(mmol·L−1) 品系排序
Line sequenceA B C D E F 平均值
MeanA B C D E F 平均值
Mean15芸芥4 15yunjie4 17 15 13 21 17 15 16.3 25 21 20 28 24 21 23 1 11芸芥32-1 11 yunjie32-1 16 13 14 13 14 10 13.3 22 18 19 19 20 15 18.8 3 16芸芥9 16yunjie9 15 14 14 15 19 12 14.8 22 19 20 22 26 18 21 2 平均值 Mean 16 14 13.7 16.3 16.7 12.3 14.8 23 19.3 19.7 23 23.3 18 21 指标排序 Index sequence 3 4 5 2 1 6 2 4 3 2 1 5 3. 讨论与结论
3.1 3个芸芥品系在4种单盐胁迫下的耐盐性差异
研究结果表明,同一个芸芥品系在某一种单盐胁迫下,发芽率等6个指标的耐盐极限值和耐盐临界值差异均较大。同一品系同一指标的耐盐临界值与耐盐极限值也不完全成正比,说明选择不同指标对3个材料种子萌发期耐盐性强弱评价结果并不完全一致。另外,植物在盐胁迫下表现出的耐盐能力大小是多种代谢的综合表现。因此,不能根据单一指标来评价植物的耐盐性强弱,应用多个指标综合评价分析[16-17]。本研究对发芽势等6个指标的耐盐临界值和极限值进行了综合分析,明确了萌发期3个芸芥品系在4种单盐胁迫下的耐盐性差异,耐NaCl和Na2SO4胁迫次序一致,即15芸芥4>11芸芥32-1>16芸芥9,耐NaHCO3胁迫依次为:16芸芥9>15芸芥4>11芸芥32-1,耐Na2CO3胁迫依次为:15芸芥4>16芸芥9>11芸芥32-1。
3.2 芸芥胚根耐盐性的适应浓度因盐碱种类不同而异
在4种盐胁迫下,3个品系胚根长的耐盐临界值和耐盐极限值均最小,说明芽期种子胚根是一个对盐分敏感的部位[15],今后在鉴定筛选耐盐种质或品种时,以胚根长作为鉴定指标。植物种子在不同的盐浓度下生长都会受到不同程度的影响[17],不同浓度盐溶液对各芸芥品系种子的发芽率、胚根长度的影响表明:在150 mmol·L−1 NaCl、90 mmol·L−1 Na2SO4、30 mmol·L−1 NaHCO3和15 mmol·L−1 Na2CO3处理下,发芽率和发芽势在参试品系间差异很大,说明这4个盐溶液适宜作为芸芥萌发期耐盐性鉴定的盐溶液浓度。
3.3 Na2SO4对芸芥种子萌发的抑制效应强于NaCl
NaCl和Na2SO4对种子萌发和幼苗生长的抑制效应主要来自Na+毒害。在Na+浓度相同时,NaCl对野生早熟禾种子萌发的影响大于Na2SO4,白小明等[15]认为这是由于NaCl比Na2SO4分子量低,容易透过种皮进入种子,使种子Na+的吸收较Na2SO4的高。研究者在对甜椒(Capsicum annuum L.)、紫花苜蓿(Medicago sativa)、高粱(Sorghum bicolor)、芦苇(Phragmites australis)、冰草(Agropyron cristatum)的研究中均发现了NaCl胁迫的生长抑制效应强于Na2SO4胁迫的类似结果[18]。与此相反,在相同Na+浓度条件下,对偃伏梾木(Cornus stolonifera)、罗勒(Ocimum basilicum)、茄属(Solanum)6种野生马铃薯和盐碱凤毛菊(Saussurea runcinata)研究的结果均证明,Na2SO4胁迫对植株生长的抑制作用强于NaCl胁迫。本试验也发现,NaCl胁迫下,芸芥发芽率、发芽指数、胚根长和胚轴长及其耐盐临界值均比Na2SO4胁迫下的高,表明Na2SO4对芸芥种子萌发的抑制效应强于NaCl胁迫。出现这种结果,可能是因为芸芥种子及幼苗Na+贮藏能力或Na+外排效率随外界溶液中阴离子种类的不同而发生变化,其他原因有待进一步探索。因此在种子萌发期,野生早熟禾等草本类和芝麻菜属作物响应NaCl和Na2SO4胁迫的特性具有明显的差异。
3.4 Na2CO3对芸芥种子萌发的抑制效应强于NaHCO3
盐害条件下植物不仅会受到渗透胁迫、离子毒害以及营养亏缺等方面的伤害[19],还会受到特有的胁变因素,如高pH胁迫等[20]。本试验结果发现,在Na+浓度相同时(30 mmol·L−1),较Na2CO3胁迫而言,NaHCO3胁迫下发芽率、发芽势、发芽指数和胚根长的耐盐临界值均较高,这表明NaHCO3对芸芥种子萌发的影响较小。这是因为Na2CO3溶液中pH值较高,对发芽势、发芽指数、发芽率、胚轴长和胚根长的抑制作用越大,即高pH的抑制作用大于Na+的胁迫效应[21]。
3.5 碱性盐或中性盐对芸芥种子萌发及幼苗生长抑制明显
有研究表明,低浓度盐胁迫能刺激植物的呼吸酶,有利于细胞膜渗透调节,从而促进种子萌发、幼苗和胚根生长发育[22-23];也有研究发现,高、低浓度盐胁迫对种子萌发、幼苗生长发育都可产生抑制作用[16, 24]。大多数研究结果表明,高浓度盐胁迫明显抑制种子萌发,幼苗不能正常生长[ 22]。本研究结果表明,低浓度盐胁迫对3个芸芥品系种子萌发没有促进作用,当碱性盐(Na+≥30 mmol·L−1)或中性盐(Na+≥180 mmol·L−1)时,芸芥种子萌发及幼苗生长受到明显抑制作用。
本研究结果表明Na2SO4对芸芥种子萌发的抑制效应强于NaCl,Na2CO3对芸芥种子萌发的抑制效应强于NaHCO3;芸芥耐NaCl和Na2SO4胁迫次序一致,均表现为15芸芥4>11芸芥32-1>16芸芥9,耐NaHCO3胁迫依次为16芸芥9>15芸芥4>11芸芥32-1,耐Na2CO3胁迫依次为15芸芥4>16芸芥9>11芸芥32-1。今后将进一步开展混合盐碱的胁迫试验,以期为芸芥选择适宜的种植土壤提供理论依据。
-
![]()
图 1 pET-28a-CD25重组质粒的构建
A:CD25基因的扩增;M:DNA Marker-2000;1∶CD25基因。B:pET-28a-CD25重组质粒的双酶切鉴定;M:DNA Marker-5000;1∶pET-28a-CD25。
Figure 1. Construction of pET-28a-CD25 recombinant plasmid
A: amplification of CD25; M: DNA marker-2000; 1: CD25. B: identification of recombinant plasmid of pET-28a-CD25 by double enzyme digestion; M: DNA marker-5000; 1: pET-28a-CD25.
![]()
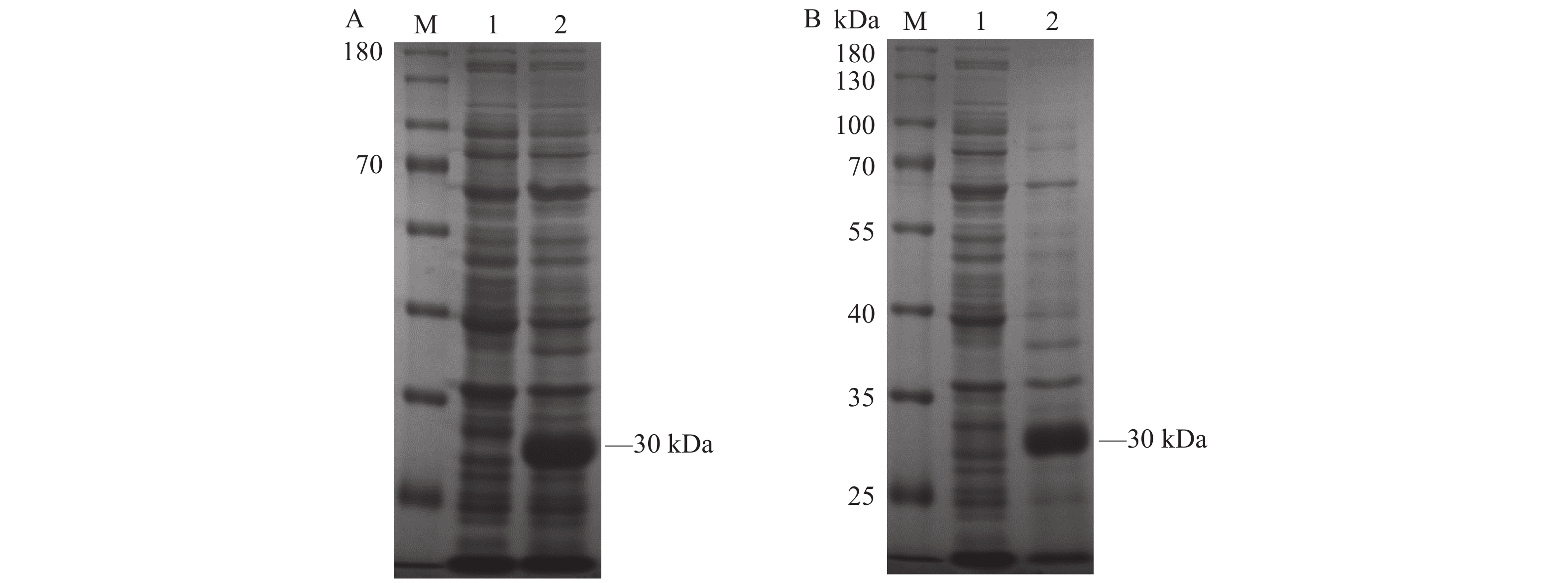
图 2 重组蛋白的诱导表达和可溶性分析
A:6×His-CD25重组蛋白SDS-PAGE分析结果;M:蛋白Marker;1:空载体pET28a(+) 诱导后的全菌体蛋白;2:E.coli Transetta (DE3)-pET28a-CD25诱导后的全菌体蛋白。 B:6×His-CD25重组蛋白可溶性鉴定;M:蛋白Marker;1:E.coli Transetta (DE3)-pET28a-CD25超声破碎后的上清;2:E.coli Transetta (DE3)-pET28a-CD25超声破碎后的沉淀。
Figure 2. Induced expression and solubility analysis of recombinant protein
A: SDS-PAGE analysis on 6×His-CD25 recombinant protein; M: protein marker; 1: whole cell protein induced by empty carrier pET-28a(+); 2: whole cell protein induced by E.coliTransetta (DE3)-pET28a-CD25; B: solubility identification of 6×His-CD25 recombinant protein; M: protein marker; 1: supernatant of E.coli Transetta (DE3)-pET28a-CD25 after ultrasonic crushing; 2: E.coli Transetta (DE3)-pET28a-CD25 precipitate after ultrasonic crushing.
![]()
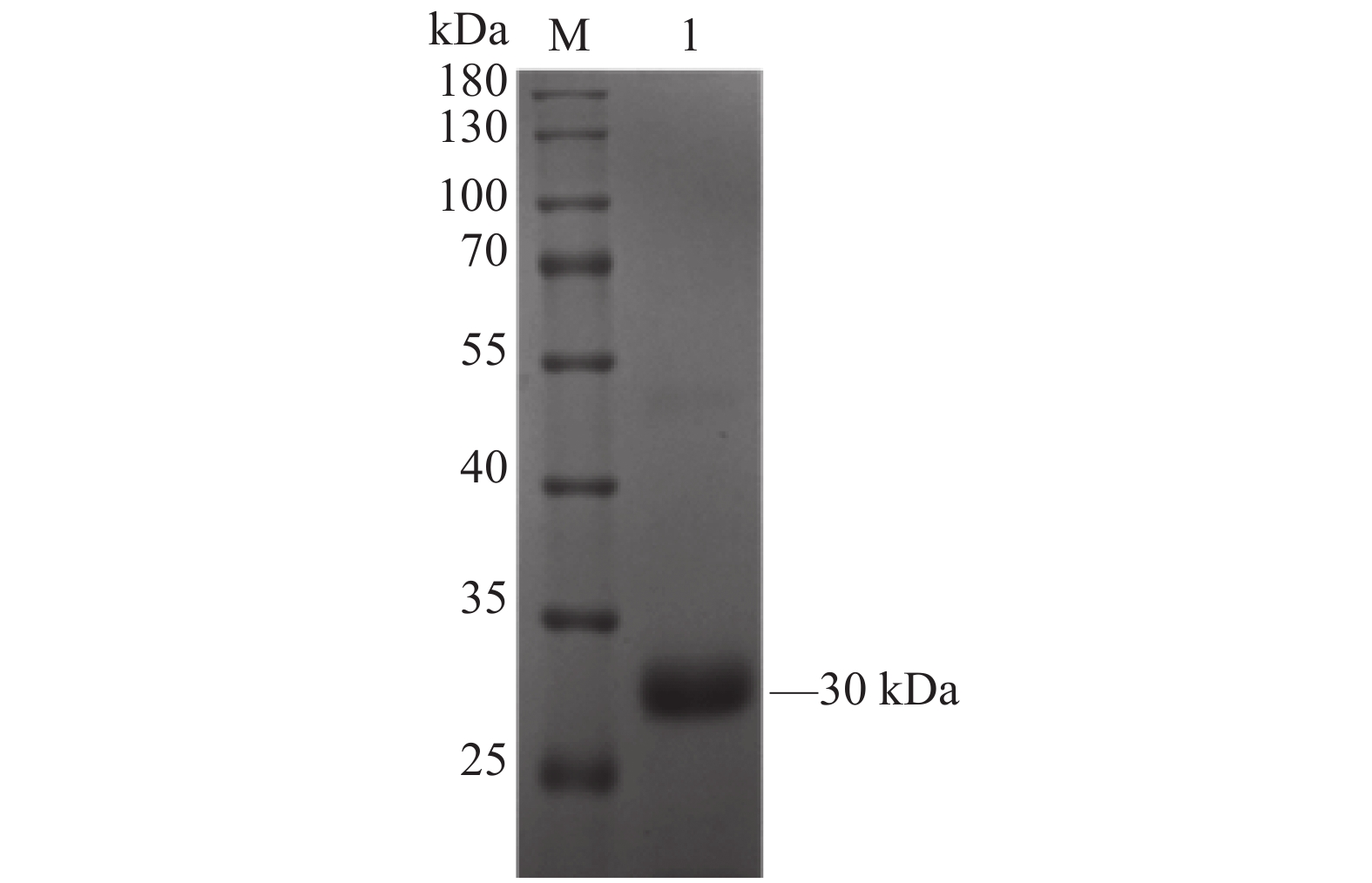
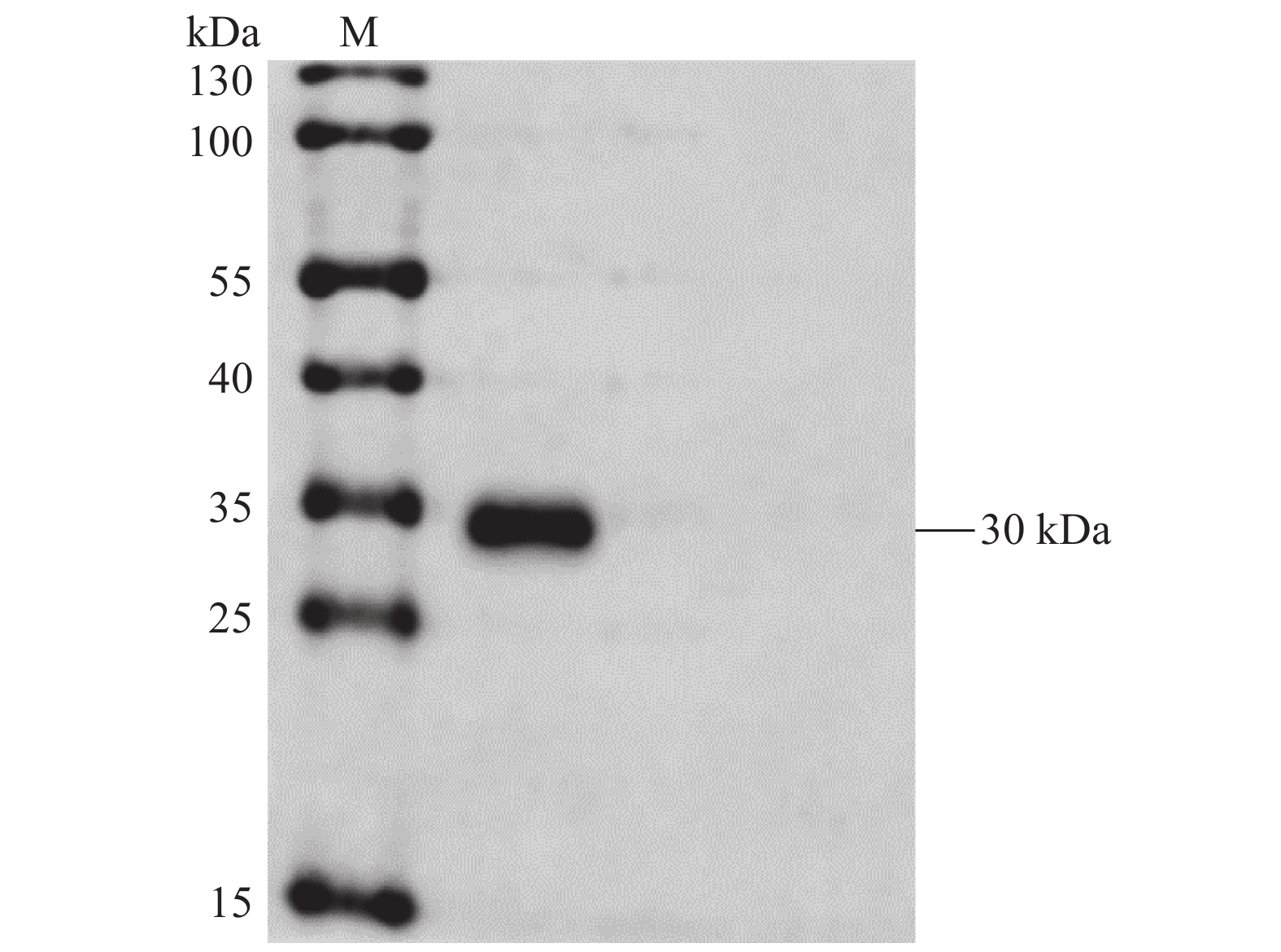
图 3 6×His-CD25重组蛋白的纯化结果
M:蛋白Marker;1:纯化后的6×His-CD25重组蛋白。
Figure 3. Purified 6×His-CD25 recombinant protein
M: Protein marker; 1: purified 6×His-CD25 recombinant protein.
![]()
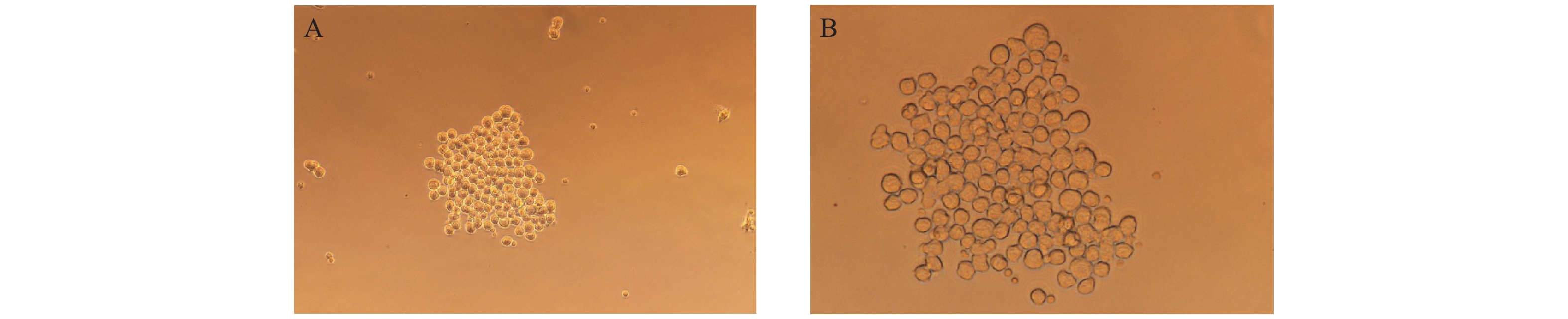
图 4 杂交瘤细胞形态
A:杂交瘤细胞(200×);B:杂交瘤细胞(400×)。
Figure 4. Morphology of hybridoma cells
A: hybridoma cells enlarged by 200×; B: hybridoma cells enlarged by 400×.
![]()
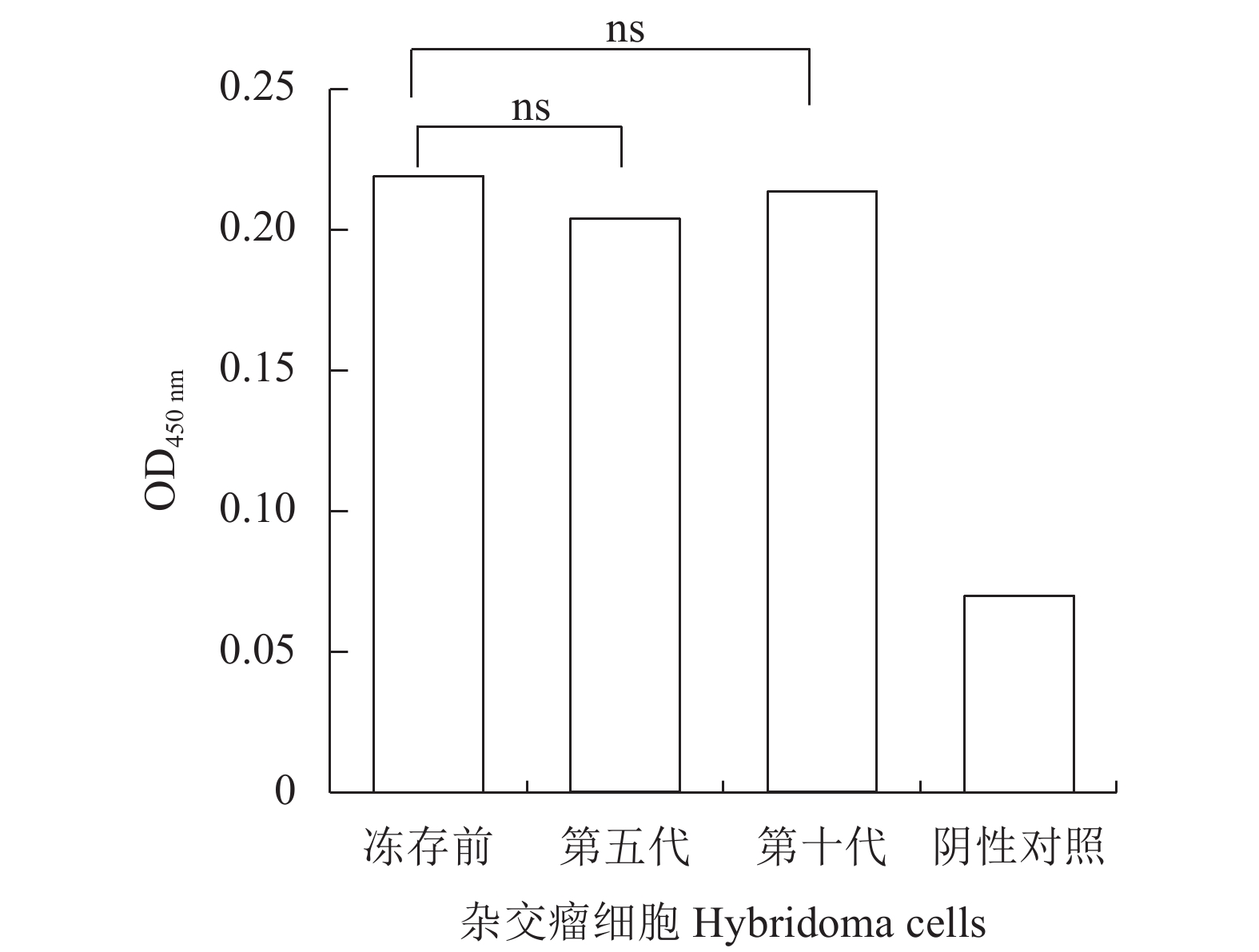
图 5 杂交瘤细胞的稳定性分析
ns表示差异不显著。
Figure 5. Stability of hybridoma cells
ns means no significant difference.
![]()
图 6 Western blotting检测单克隆抗体的特异性结果
M:蛋白Marker;1:CD25表达菌株诱导后全菌体蛋白;2:其他6×His标签蛋白;3:BSA(牛血清白蛋白)。
Figure 6. Specificity of monoclonal antibody tested by western blotting
M: protein marker; 1: whole cell protein induced by CD25 expression strain; 2. other 6×His tag proteins; 3: BSA (bovine serum albumin).
![]()
图 7 单克隆抗体与CD25蛋白的间接免疫荧光反应(100×)
A:转染pEΔgfp-CD25的293细胞; B:阴性对照。
Figure 7. Indirect immunofluorescence reaction between monoclonal antibody and CD25 protein(100×)
A: 293 cells transfected with pEΔgfp-CD25; B: negative control.
![]()
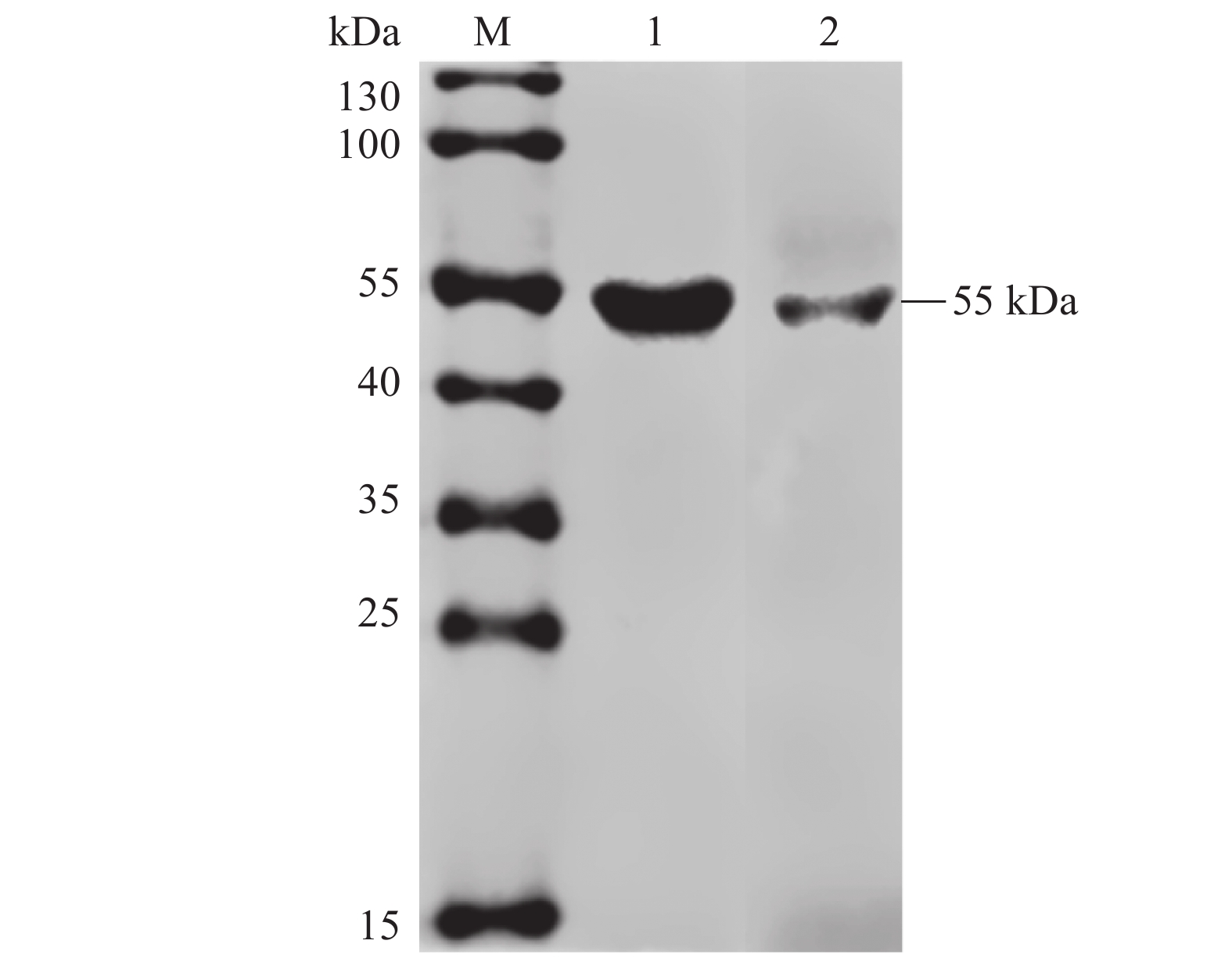
图 8 单克隆抗体的Western-blotting鉴定
M:蛋白Marker;1∶猪脾脏组织蛋白样;2:猪肺脏组织蛋白样。
Figure 8. Western blot identification on monoclonal antibody
M: Protein marker; 1: pig spleen tissue protein specimen; 2. pig lung tissue protein specimen.
表 1 6×His-CD25重组蛋白不同抗原包被浓度和血清稀释倍数下的OD450值
Table 1 OD450 under optimum antigen coating concentration and serum dilution multiple of 6×His-CD25 recombinant protein
血清类型
Serum type稀释倍数
Dilution times抗原包被浓度
Antigen coating concentration250/(ng·mL−1) 125/(ng·mL−1) 62.5/(ng·mL−1) 31.25/(ng·mL−1) 15.63/(ng·mL−1) 7.81/(ng·mL−1) 3.91/(ng·mL−1) 1.95/(ng·mL−1) 阳性血清稀释倍数
Positive serum dilution1∶ 2000 OVER OVER OVER OVER 3.317 3.100 2.977 2.790 1∶ 4000 OVER OVER OVER 2.573 1.873 1.723 1.417 1.227 1∶ 8000 OVER OVER 2.556 1.981 1.583 1.124 0.925 0.810 1∶ 16000 OVER 2.661 1.854 1.068 0.692 0.352 0.252 0.265 1∶ 32000 2.221 1.622 0.826 0.444 0.189 0.137 0.129 0.147 1∶ 64000 1.569 0.871 0.356 0.152 0.125 0.108 0.086 0.123 阴性血清
Negative serum0.060 0.077 0.076 0.078 0.079 0.069 0.056 0.056 OVER表示OD450处的吸光度大于4.0。表2同。
OVER: OD450 greater than 4.0. Same for Table 2. 下载: 导出CSV
下载: 导出CSV
表 2 杂交瘤细胞冻存前后的培养液上清抗体效价
Table 2 Antibody titer of hybridoma cell supernatant before and after cryopreservation
杂交瘤细胞
Hybridoma cells上清稀释倍数
Supernatant dilution factor1∶ 1000 1∶ 2000 1∶ 4000 1∶ 8000 1∶ 16000 1∶ 32000 1∶ 64000 1∶ 128000 冻存前
Before cryopreservationOVER 3.729 2.787 1.725 0.933 0.457 0.216 0.096 复苏后
After resuscitationOVER 2.912 2.188 1.287 0.742 0.293 0.177 0.087 阴性对照
Negative control0.049 0.049 0.051 0.050 0.051 0.050 0.049 0.051
下载: 导出CSV
表 3 小鼠腹水单克隆抗体效价
Table 3 Monoclonal antibody titer of mouse ascites
诱导佐剂
Adjucant type小鼠腹水稀释倍数
Ascites dilution factor in mice1∶ 128000 1∶ 256000 1∶ 512000 1∶ 1024000 1∶ 2048000 1∶ 4096000 弗氏不
完全佐剂
Freund's incomplete adjuvant3.670 3.279 2.444 1.356 0.722 0.256 液体石蜡
Liquid paraffin2.552 1.539 0.899 0.409 0.209 0.090 阴性对照
Negative control0.046 0.046 0.047 0.046 0.047 0.050
下载: 导出CSV
-
[1] LUO H, ZHU Y, GUO B, et al. Causal relationships between CD25 on immune cells and hip osteoarthritis [J]. Frontiers in Immunology, 2023, 14: 1247710. DOI: 10.3389/fimmu.2023.1247710
[2] FONTENOT J D, RASMUSSEN J P, GAVIN M A, et al. A function for interleukin 2 in Foxp3-expressing regulatory T cells [J]. Nature Immunology, 2005, 6(11): 1142−1151. DOI: 10.1038/ni1263
[3] SAKAGUCHI S, SAKAGUCHI N, ASANO M, et al. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases [J]. Journal of Immunology (Baltimore, Md. : 1950), 1995, 155(3): 1151−1164. DOI: 10.4049/jimmunol.155.3.1151
[4] 刘莉, 姚军霞, 丁乾, 等. 非霍奇金淋巴瘤患者外周血CD4+CD25high调节性T细胞研究 [J]. 中国实验血液学杂志, 2006, 14(1):119−122. DOI: 10.3969/j.issn.1009-2137.2006.01.027 LIU L, YAO J X, DING Q, et al. CD4+ CD25high regulatory T cells in peripheral blood of patients with B cell non-Hodgkin’s lymphoma [J]. Journal of Experimental Hematology, 2006, 14(1): 119−122. (in Chinese) DOI: 10.3969/j.issn.1009-2137.2006.01.027
[5] HAN Y K, YU C C, YU Y, et al. CD25+ B cells produced IL-35 and alleviated local inflammation during experimental periodontitis [J]. Oral Diseases, 2022, 28(8): 2248−2257. DOI: 10.1111/odi.13939
[6] 安田丽, 李亮亮, 赵丽. 骨髓微环境影响调节性T细胞促进血液恶性肿瘤发展的研究进展 [J]. 中国免疫学杂志, 2021, 37(5):635−642. DOI: 10.3969/j.issn.1000-484X.2021.05.024 AN T L, LI L L, ZHAO L. Progress on influence of bone marrow microenvironment on regulatory T cells promoting development of hematological malignancies [J]. Chinese Journal of Immunology, 2021, 37(5): 635−642. (in Chinese) DOI: 10.3969/j.issn.1000-484X.2021.05.024
[7] TAGUCHI O, TAKAHASHI T. Administration of anti-interleukin-2 receptor alpha antibody in vivo induces localized autoimmune disease [J]. European Journal of Immunology, 1996, 26(7): 1608−1612. DOI: 10.1002/eji.1830260730
[8] MCHUGH R S, SHEVACH E M. Cutting edge: Depletion of CD4+CD25+ regulatory T cells is necessary, but not sufficient, for induction of organ-specific autoimmune disease [J]. Journal of Immunology, 2002, 168(12): 5979−5983. DOI: 10.4049/jimmunol.168.12.5979
[9] LOWENTHAL J W, ZUBLER R H, NABHOLZ M, et al. Similarities between interleukin-2 receptor number and affinity on activated B and T lymphocytes [J]. Nature, 1985, 315(6021): 669−672. DOI: 10.1038/315669a0
[10] HUSS D J, PELLERIN A F, COLLETTE B P, et al. Anti-CD25 monoclonal antibody Fc variants differentially impact regulatory T cells and immune homeostasis [J]. Immunology, 2016, 148(3): 276−286. DOI: 10.1111/imm.12609
[11] FLYNN M J, HARTLEY J A. The emerging role of anti-CD25 directed therapies as both immune modulators and targeted agents in cancer [J]. British Journal of Haematology, 2017, 179(1): 20−35. DOI: 10.1111/bjh.14770
[12] AMBROSETTI A, NADALI G, VINANTE F, et al. Soluble interleukin-2 receptor in hairy-cell leukemia: A reliable marker of disease [J]. International Journal of Clinical & Laboratory Research, 1993, 23(1): 34−37.
[13] DAMOISEAUX J. The IL-2 - IL-2 receptor pathway in health and disease: The role of the soluble IL-2 receptor [J]. Clinical Immunology, 2020, 218: 108515. DOI: 10.1016/j.clim.2020.108515
[14] ZHOU M, FANG H S, DU M, et al. The modulation of regulatory T cells via HMGB1/PTEN/β-catenin axis in LPS induced acute lung injury [J]. Frontiers in Immunology, 2019, 10: 1612. DOI: 10.3389/fimmu.2019.01612
[15] NIXON D F, AANDAHL E M, MICHAËLSSON J. CD4+CD25+ regulatory T cells in HIV infection [J]. Microbes and Infection, 2005, 7(7/8): 1063−1065.
[16] LUO Y, XUE Y, MAO L Y, et al. Activation phenotype of Mycobacterium tuberculosis-specific CD4+ T cells promoting the discrimination between active tuberculosis and latent tuberculosis infection [J]. Frontiers in Immunology, 2021, 12: 721013. DOI: 10.3389/fimmu.2021.721013
[17] WANG L S, CHOW K C, LI W Y, et al. Clinical significance of serum soluble interleukin 2 receptor-alpha in esophageal squamous cell carcinoma [J]. Clinical Cancer Research, 2000, 6(4): 1445−1451.
[18] NAKASE K, TSUJI K, TAMAKI S, et al. Elevated levels of soluble interleukin-2 receptor in serum of patients with hematological or non-hematological malignancies [J]. Cancer Detection and Prevention, 2005, 29(3): 256−259. DOI: 10.1016/j.cdp.2005.03.001
[19] ATANACKOVIC D, CAO Y R, LUETKENS T, et al. CD4+CD25+FOXP3+ T regulatory cells reconstitute and accumulate in the bone marrow of patients with multiple myeloma following allogeneic stem cell transplantation [J]. Haematologica, 2008, 93(3): 423−430. DOI: 10.3324/haematol.11897
[20] GUALCO G, CHIOATO L, WEISS L M, et al. Analysis of human T-cell lymphotropic virus in CD25+ anaplastic large cell lymphoma in children [J]. American Journal of Clinical Pathology, 2009, 132(1): 28−33. DOI: 10.1309/AJCP6Q7QMUVGMVMF
[21] YANO T, FUKUYAMA Y, YOKOYAMA H, et al. Interleukin-2 receptors in pulmonary adenocarcinoma tissue [J]. Lung Cancer, 1996, 16(1): 13−19. DOI: 10.1016/S0169-5002(96)00608-3
[22] TARTOUR E, MOSSERI V, JOUFFROY T, et al. Serum soluble interleukin-2 receptor concentrations as an independent prognostic marker in head and neck cancer [J]. Lancet, 2001, 357(9264): 1263−1264. DOI: 10.1016/S0140-6736(00)04420-2
[23] SCHWARTZ M, ZHANG Y, ROSENBLATT J D. B cell regulation of the anti-tumor response and role in carcinogenesis [J]. Journal for Immunotherapy of Cancer, 2016, 4: 40. DOI: 10.1186/s40425-016-0145-x
[24] WONGYANIN P, BURANAPRADITKUN S, CHOKESHAI-USAHA K, et al. Induction of inducible CD4+CD25+Foxp3+ regulatory T lymphocytes by porcine reproductive and respiratory syndrome virus (PRRSV) [J]. Veterinary Immunology and Immunopathology, 2010, 133(2/3/4): 170−182.
[25] SILVA-CAMPA E, MATA-HARO V, MATEU E, et al. Porcine reproductive and respiratory syndrome virus induces CD4+CD8+CD25+Foxp3+ regulatory T cells (Tregs) [J]. Virology, 2012, 430(1): 73−80. DOI: 10.1016/j.virol.2012.04.009
[26] KÄSER T, GERNER W, HAMMER S E, et al. Phenotypic and functional characterisation of porcine CD4+CD25(high) regulatory T cells [J]. Veterinary Immunology and Immunopathology, 2008, 122(1/2): 153−158.
[27] ŠTĚPÁNOVÁ K, ŠINKORA M. The expression of CD25, CD11b, SWC1, SWC7, MHC-II, and family of CD45 molecules can be used to characterize different stages of γδ T lymphocytes in pigs [J]. Developmental & Comparative Immunology, 2012, 36(4): 728−740.
[28] PICKERING B S, HARDHAM J M, SMITH G, et al. Protection against henipaviruses in swine requires both, cell-mediated and humoral immune response [J]. Vaccine, 2016, 34(40): 4777−4786. DOI: 10.1016/j.vaccine.2016.08.028
[29] SURADHAT S, SADA W, BURANAPRADITKUN S, et al. The kinetics of cytokine production and CD25 expression by porcine lymphocyte subpopulations following exposure to classical swine fever virus (CSFV) [J]. Veterinary Immunology and Immunopathology, 2005, 106(3/4): 197−208.
[30] LOWENTHAL J W, CORTHÉSY P, TOUGNE C, et al. High and low affinity IL 2 receptors: Analysis by IL 2 dissociation rate and reactivity with monoclonal anti-receptor antibody PC61 [J]. Journal of Immunology, 1985, 135(6): 3988−3994. DOI: 10.4049/jimmunol.135.6.3988
[31] ALLOUCHE M, SAHRAOUI Y , AUGERY-BOURGET Y, et al. Interleukin 2 receptors[J]. Leukemia research, 1990, 14(8): 699-703.
[32] LEONARD W J, DEPPER J M, CRABTREE G R, et al. Molecular cloning and expression of cDNAs for the human interleukin-2 receptor [J]. Nature, 1984, 311(5987): 626−631. DOI: 10.1038/311626a0
[33] KOKUHO T, HIRAIWA H, YASUE H, et al. Cloning and chromosomal assignment of the porcine interleukin-2 receptor alpha (IL-2Ralpha) gene [J]. Journal of Veterinary Medical Science, 2000, 62(8): 841−847. DOI: 10.1292/jvms.62.841
[34] ABRAMS V K, HWANG B, LESNIKOVA M, et al. A novel monoclonal antibody specific for canine CD25 (P4A10): Selection and evaluation of canine tregs [J]. Veterinary Immunology and Immunopathology, 2010, 135(3/4): 257−265.
[35] MALOY K J, POWRIE F. Regulatory T cells in the control of immune pathology [J]. Nature Immunology, 2001, 2(9): 816−822. DOI: 10.1038/ni0901-816
[36] 杨晓蓉, 杨启梅, 韩想利, 等. CD4+CD25+Foxp3+Treg细胞及其相关因子在梅尼埃病中的意义 [J]. 听力学及言语疾病杂志, 2022, 30(5):469−472. DOI: 10.3969/j.issn.1006-7299.2022.05.003 YANG X R, YANG Q M, HAN X L, et al. Clinical value of CD4+CD25+Foxp3+Treg cells and their related factors in Meniere’s disease [J]. Journal of Audiology and Speech Pathology, 2022, 30(5): 469−472. (in Chinese) DOI: 10.3969/j.issn.1006-7299.2022.05.003
[37] RONCAROLO M G, BATTAGLIA M. Regulatory T-cell immunotherapy for tolerance to self antigens and alloantigens in humans [J]. Nature Reviews Immunology, 2007, 7(8): 585−598. DOI: 10.1038/nri2138
[38] VOSS S D, SONDEL P M, ROBB R J. Characterization of the interleukin 2 receptors (IL-2R) expressed on human natural killer cells activated in vivo by IL-2: Association of the p64 IL-2R gamma chain with the IL-2R beta chain in functional intermediate-affinity IL-2R [J]. Journal of Experimental Medicine, 1992, 176(2): 531−541. DOI: 10.1084/jem.176.2.531
[39] MANDRIOLI L, SARLI G, ZENGARINI M, et al. Immunohistochemical MHC-II and interleukin 2-R (CD25) expression in lymph nodes of pigs with spontaneous postweaning multisystemic wasting syndrome (PMWS) [J]. Veterinary Pathology, 2006, 43(6): 993−997. DOI: 10.1354/vp.43-6-993
-
期刊类型引用(6)
1. 宋琳琳,陈红芝,毋柳柳,李畅,孟丽,孔维丽. 基于重测序的23份香菇种质资源全基因组序列分析. 中国瓜菜. 2024(03): 28-34 .  百度学术
百度学术
2. 周超,马银鹏,包旭翔,张介驰. DNA分子标记技术在黑木耳中的研究进展. 北方园艺. 2023(07): 125-131 . 百度学术
3. 吴小燕,鲍红春,李小雷,于海滨,王海霞,于传宗,庞杰,王锋. 26个香菇菌株的遗传多样性ISSR分析. 种子. 2023(05): 63-67+85 . 百度学术
4. 赵光辉,吴汉琼,方洪枫. 基于ISSR和RAPD分子标记的15株糙皮侧耳遗传多样性分析. 热带农业科学. 2023(12): 29-33 . 百度学术
5. 罗影,贾培松,努尔孜亚·亚力买买提,祁颢萱,赵振豪,贾文捷. 新疆野生中国美味蘑菇的遗传多样性分析. 新疆农业科学. 2022(11): 2707-2713 . 百度学术
6. 胡晓强,赵光辉,马玮超,李峰. 香菇栽培菌株区别性鉴定及其比较试验. 食用菌. 2020(02): 35-37 . 百度学术
其他类型引用(3)






计量
- 文章访问数: 40
- HTML全文浏览量: 12
- PDF下载量: 2
- 被引次数: 9
