

Metabolomics of Lactiplantibacillus plantarum Fermentation on Ginseng Residues
-
摘要:目的
研究植物乳杆菌N-25发酵党参渣对其代谢物成分变化及相关代谢通路的影响。
方法以植物乳杆菌N-25发酵党参渣,设置空白对照组(Y1)、低剂量植物乳杆菌N-25组(Y2,接菌量1%)、中剂量植物乳杆菌N-25组(Y3,接菌量3%)、高剂量植物乳杆菌N-25组(Y4,接菌量5%),通过非靶向代谢组学方法测定发酵前后党参渣中的物质成分变化,并对差异代谢物和代谢通路进行比较分析。
结果经植物乳杆菌N-25发酵后,党参渣的代谢物由
2710 种增加至2736 种,从中筛选到560种共有的差异代谢物,被注释的有227种,包括脂质和类脂分子(14.54%)、苯丙烷和聚酮化合物(12.33%)、莽草酸及苯丙烷类(11.89%)、有机杂环化合物(10.13%)、生物碱及衍生物(8.81%)、有机酸及衍生物(8.37%)以及脂肪酸(7.93%)等,其中黄酮类化合物含量增加1.13倍,木脂素含量增加1.04倍,生物碱及其衍生物含量增加1.01倍,有机酸及其衍生物含量增加1.16倍。通过对共有差异代谢物富集的代谢通路进行分析,得到差异最具显著的代谢通路为异喹啉生物碱生物合成通路。另外,Y2、Y3、Y4组特有的差异代谢物数量为128、132、204种。Y2组特有的差异代谢物富集到的关键通路为氰氨酸代谢通路,Y3与Y4组特有的差异代谢物富集到的关键通路为缬氨酸、亮氨酸、异亮氨酸生物合成通路。不同的接菌量可以诱导党参渣产生不同的代谢产物,且涉及的功能也有所不同。结论植物乳杆菌N-25发酵党参渣,促进了黄酮类化合物、木脂素、生物碱及其衍生物、有机酸及其衍生物等活性物质的生成与释放;其共有差异代谢物富集的显著性差异代谢通路为异喹啉生物碱生物合成。
Abstract:ObjectiveMetabolites generated in the ginseng residue fermentation with Lactiplantibacillus plantarum were analyzed.
MethodsSpent residues of ginseng (Codonopsis pilosula) were inoculated with L. plantarum N-25 at low-dose of 1% (Y2), medium-dose of 3% (Y3) or high-dose of 5% (Y4) along with control of no inoculation (Y1). Non-targeted metabolomics was conducted to study the changes in the metabolites, and analyses of differential metabolites and metabolic pathways performed on the solid medium during fermentation.
ResultsThe fermentation raised the number of differential metabolites in the ginseng substrate from
2710 to2735 . Among them, 560 were found commonly existing in all samples, while 128 identified as unique in Y2, 132 in Y3, and 204 in Y4. And 227 metabolites were annotated which included lipids and lipoid molecules (14.54%), phenylpropanoids and polyketide compounds (12.33%), shikimate and phenylpropanoid (11.89%), organic heterocycle compounds (10.13%), biogenic amines and derivatives (8.81%), organic acids and derivatives (8.37%), and fatty acids (7.93%). Contents of some of the substances increased significantly in the fermentation. For instance, flavonoids rose by 1.13 times, lignans by 1.04 times, alkaloids and derivatives by 1.01 times, and organic acids and derivatives by 1.16 times. Of the metabolic pathways, that of isoquinoline alkaloid biosynthesis pathways was the most significantly enriched by the fermentation. The key pathway enhanced by the unique metabolites in Y2 was associated with the cyanogenic amino acid metabolism, while those in Y3 and Y4 with valine, leucine, and isoleucine biosynthesis. The inoculation dosage of L. plantarum N-25 significantly altered the contents and functions of the metabolites generated in the fermentation.ConclusionThe fermentation of C. pilosula residues inoculated with L. plantarum N-25 released functional ingredients such as flavonoids, lignans, alkaloids and derivatives, and organic acids and derivatives. It significantly enriched the pathway of isoquinoline alkaloid biosynthesis.
-
0. 引言
【研究意义】党参(Codonopsis pilosula)为桔梗科(Campanulaceae)党参属(Codonopsis)植物,其干燥根性味甘平,归脾、肺经,具有健脾益肺、养血生津的作用[1],现代药理研究表明党参具有促进造血性能、调节血糖、抗炎、延缓衰老、抗缺氧和加强机体免疫力等多种功效[2]。目前,党参的年消耗量在5万t左右[3],其用量持续增加导致党参渣的大量产出,由于提取方式和加工工艺制约,党参渣无法得到很好利用,而直接废弃则会造成较大的资源浪费[4]。【前人研究进展】已有研究将党参渣用于饲料添加剂,可促进麻鸡生产性能的改善,并增加麻鸡养殖的经济效益[5];除此之外,有研究报道,废弃药渣(包括党参渣)可通过生物转化技术用于生产沼气,或利用热化学转化技术将其转换成燃料物质[6],或可通过微生物技术生产包含多种活性成分的制剂或新药[7]。因此,开展党参渣的资源性物质二次开发利用,提高党参渣利用率,促进党参产业持续高速发展,成为当前党参产业健康发展的重要一环[8]。近年来,随着国家减抗和替抗政策的陆续出台,微生物发酵中药渣的工艺在畜牧业领域的应用受到广为关注[9]。孟晓燕[10]经研究发现,发酵中药渣能提高牛血清中过氧化物歧化酶、血清总蛋白、谷胱甘肽和免疫球蛋白等血清指标,提升抗氧化能力。李华伟[11]经研究发现,饲喂3%发酵中药渣(当归Angelicae Sinensis Radix、黄芪Astragalus membranaceus、白芍Cynanchum otophyllum和熟地黄Rehmannia glutinosa)可增加断奶仔猪的平均日增重,提高生产性能。植物乳杆菌(Lactiplantibacillus plantarum)是一种重要的益生菌,具有改善肠道菌群功能、抑制有害微生物的作用,近几年被广泛应用于中药发酵[12−15],其产生的胞外多糖具有抗氧化活性[16]。Fu等[17]利用植物乳杆菌发酵中药材红景天,发现其抗氧化和抗光老化活性均明显提高。Eweys等[18]研究发现,利用植物乳杆菌发酵肉桂,不但能提高肉桂的抗氧化活性,而且肉桂中的没食子酸、对羟基苯甲酸、儿茶素和绿原酸等活性成分也有一定程度的增加。【本研究切入点】党参渣利用微生物技术处理是目前较常见的方式,但目前较少经植物乳杆菌进行发酵二次利用的报道[19−20]。【拟解决的关键问题】本研究首次采用植物乳杆菌发酵党参渣,并将代谢组学的方法应用于植物乳杆菌发酵党参渣的研究,分析经植物乳杆菌N-25发酵后党参渣中的代谢物成分以及代谢通路方面的变化,以期揭示植物乳杆菌N-25发酵对党参渣代谢的影响,为党参渣资源化利用提供科学依据。
1. 材料与方法
1.1 试验材料
1.1.1 党参渣及植物乳杆菌
党参渣(醇提来源,后经去醇、干燥)回收自福建九为生物科技有限公司。植物乳杆菌N-25(N-25)分离自宁夏泡菜,保存于本实验室。在前期研究发现,N-25菌株具有良好的生物学活性,用该菌株发酵饲料可释放更多的不饱和脂肪酸和必需氨基酸等物质,促进营养物质的消化吸收和改善肠道健康[21]。
1.1.2 试剂及仪器
甲醇(LC-MS级,CNW Technologies,上海),乙腈( LC-MS级,CNW Technologies,上海),超纯水(屈臣氏),乙酸(LC-MS级,SIGMA-ALDRICH,德国),异丙醇(LC-MS级,CNW Technologies,上海),超高效液相色谱仪(Vanquish,Thermo Fisher Scientific,美国),高分辨质谱仪(Orbitrap Exploris 120,Thermo Fisher Scientific,美国),离心机(Heraeus Fresco17,Thermo Fisher Scientific,美国),天平(BSA124S-CW,Sartorius,德国),超声仪(PS-60AL,雷德邦电子有限公司,深圳),匀浆机(JXFSTPRP-24,净信科技有限公司,上海),冷冻干燥机(LGJ-10C,四环福瑞科仪科技发展有限公司,北京)。
1.2 试验方法
1.2.1 植物乳杆菌的增殖
在烧杯中加入脑心浸出液肉汤(BHI)培养基38.5 g,加入
1000 mL蒸馏水后加热,冷却后于121 ℃高压灭菌15 min,放凉备用。冰箱内取出4 ℃保存的植物乳杆菌,以1∶10的比例加入以上BHI液体培养基中,充分摇匀,在35 ℃、100 r·min−1条件下于恒温摇床中培养24 h后,4 ℃冰箱存放备用。取扩增好的菌液作10倍递增稀释,得到菌液浓度为1×108 CFU·mL−1。1.2.2 党参渣发酵
称取干燥后的党参渣100.00 g于锥形瓶内,加入一定比例蒸馏水,于121 ℃高压蒸汽灭菌锅内灭菌20 min,待冷却至室温后,于超净工作台内接种1×108 CFU·mL−1 N-25,摇匀后放入恒温培养箱。参照秦楠等的植物乳杆菌发酵工艺(温度31 ~39 ℃、发酵时间2 ~10 d、接菌量0.5% ~4%、料液比12.5% ~25%)[22],以黄酮含量作为标准观察发酵效果,据此确定本研究的发酵工艺:发酵温度35 ℃,发酵时间2 d,含水量30%进行有氧发酵。无菌水作为空白对照组(Y1),低剂量N-25组(Y2)设置1%(菌液含量0.3×108 CFU·mL−1)接菌量;中剂量N-25组(Y3)设置3%(菌液含量0.9×108 CFU·mL−1)接菌量;高剂量N-25组(Y4)设置5%(菌液含量1.5×108 CFU·mL−1)接菌量。
1.2.3 党参渣发酵代谢物的提取
试验样本送至上海百趣生物医学科技有限公司进行提取并分析。将样品冷冻干燥后研磨,低温称取25 mg样品于EP管中,加入匀浆珠,后加入
1000 μL提取液(甲醇∶乙腈∶水=2∶2∶1,V/V,含同位素标记内标混合物);涡旋混匀30 s;放入匀浆仪中匀浆(35 Hz,4 min),再转移至冰水浴超声5 min,此步骤重复3次;−40 ℃静置1 h;将样品4 ℃、12000 r·min−1,离心15 min,取上清液;上清液于4 ℃、12000 r·min−1,离心15 min;取上清液上机检测;所有样品另取等量上清液混合成QC样品上机检测。1.2.4 上机检测
针对发酵中药有较多的非极性代谢物,本试验使用Vanquish超高效液相色谱仪,通过Phenomenex Kinetex C18(2.1 mm×50 mm,2.6 μm)液相色谱柱对目标化合物进行色谱分离,条件为:液相色谱A相为水相,含0.01%乙酸,B相为异丙醇∶乙腈(1∶1,V/V)。样品盘温度:4 ℃,进样体积:2 μL。使用Xcalibur软件分析一级、二级质谱数据。
1.3 数据统计与分析
原始数据经ProteoWizard软件转成mzXML格式后,使用R包进行代谢物鉴定,所用数据库为BiotreeDB(V3.0),用R包进行可视化,采用Ropls进行主成分分析(principal component analysis, PCA)、正交偏最小二乘判别分析(orthogonal partialleast squares discriminant analysis, OPLS-DA);采用MetaboAnalyst对差异代谢物进行京都基因与基因组百科全书(Kyoto encyclopediaof genes and genomes, KEGG)代谢通路富集分析。基于SPSS26.0对相关组学数据进行单因素方差分析及差异显著性检验,使用Adobe Photoshop 2022进行绘图。
2. 结果与分析
2.1 质控分析
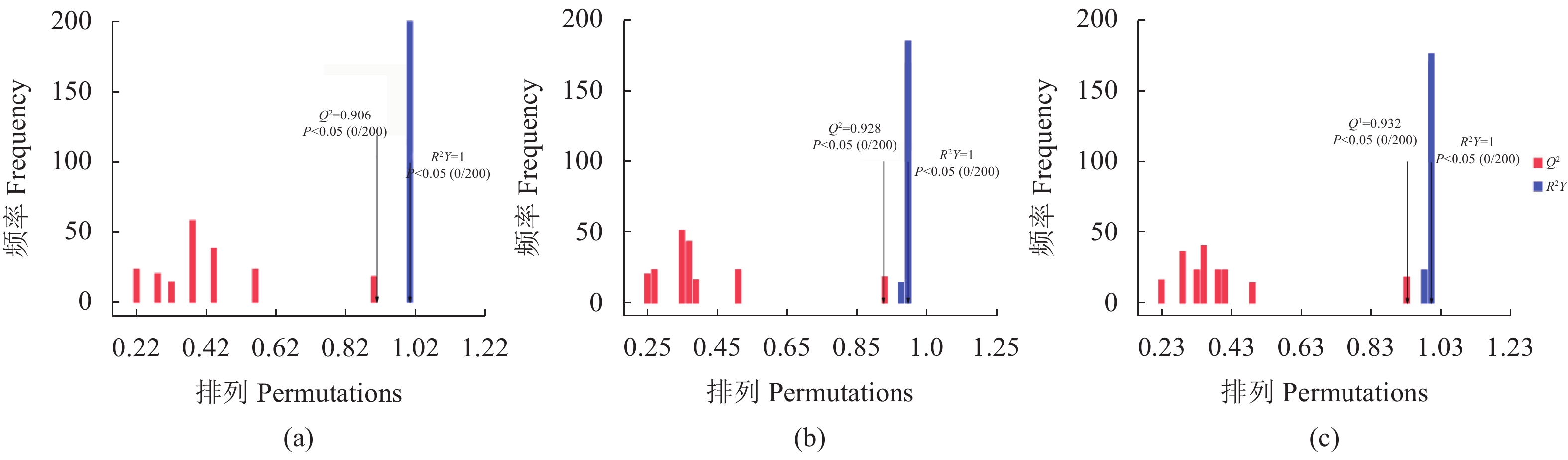
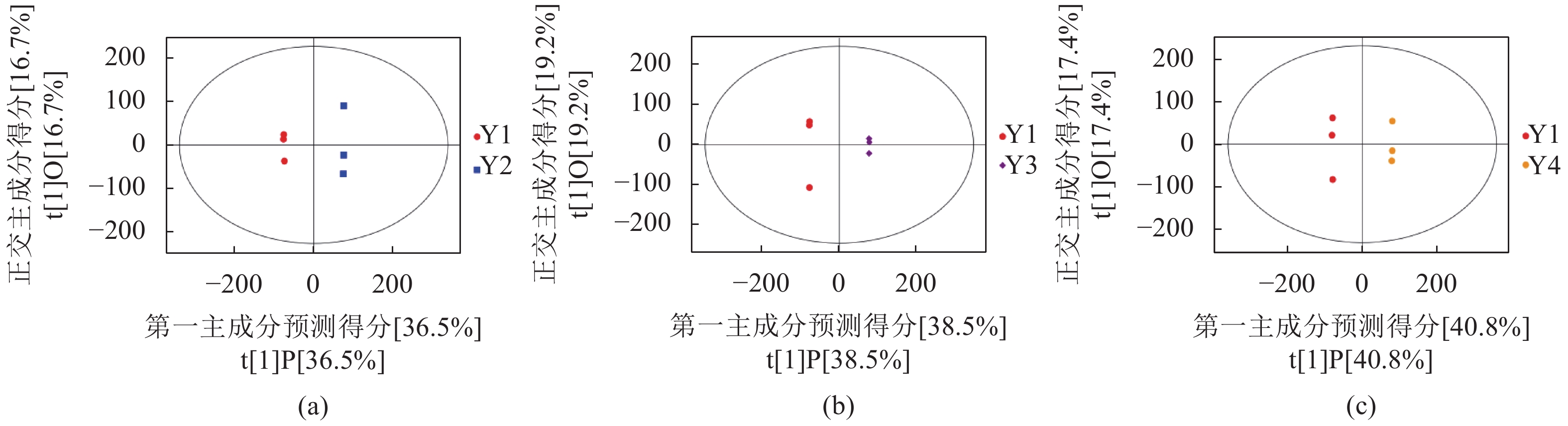
从图1中可知试验组(Y2、Y3和Y4)与Y1在OPLS-DA得分图上样本间离散程度不明显,均匀地分布于PC1左右两侧,说明组间差异小,组间整体代谢水平差异较大。通过随机改变分类变量Y的排列顺序,多次(次数n=200)建立对应的OPLS-DA模型以获取随机模型的R和Q值,来防止OPLS-DA模型过拟合并考察模型的质量,由图2可知Y1 vs. Y2的R2截距为0.99,Q2截距为−0.04;Y1 vs. Y2的R2截距为0.99,Q2截距为−0.09;Y1 vs. Y3的R2截距为0.99,Q2截距为−0.12,说明这3组OPLS-DA模型可靠。因此后续差异代谢物筛选可依据OPLS-DA模型。
![]() 图 2 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的OPLS-DA模型得分(a)Y1vs.Y2,(b)Y1vs.Y3,(c)Y1vs.Y4。Figure 2. OPLS-DA model scores for Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.
图 2 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的OPLS-DA模型得分(a)Y1vs.Y2,(b)Y1vs.Y3,(c)Y1vs.Y4。Figure 2. OPLS-DA model scores for Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.2.2 代谢物分类统计
原始数据包含3个质控样本和12个试验样本。Y1组中鉴定到
2710 个代谢物,经N-25发酵后,试验组共鉴定到2736 种代谢物。发酵产物中的代谢物质如表1所示,其中主要的代谢物有脂类和类脂分子(包括柠檬酸、棕榈酸、月桂酸、γ-亚麻酸、黄芪皂苷Ⅲ、亚麻酸、丙戊酸、亚油酸、黄芪皂苷Ⅱ等代谢物),占比4.349%;莽草酸和苯丙烷类(包括异银杏双黄酮、去甲基川陈皮素、桑黄酮 E等黄酮类物质;阿魏内酯A、佛手苷内酯、异紫花前胡内酯等香豆素类代谢物;咖啡酸、肉桂酸苄酯、柠檬酸B、荜澄茄脂酮等木脂素及其他代谢物),占比4.094%;苯丙烷类和聚酮(包括川陈皮素、黄芩苷、香叶木素、7,8-二羟基黄酮等黄酮类物质;异黄酮类及其他代谢物),占比3.509%;有机杂环化合物(包括吡啶及衍生物、吲哚和衍生物、哌啶、吡喃、呋喃并吡喃类、喹啉及衍生物、吲哚和衍生物等物质),占比3.253%;苯类化合物(包括异香兰素、4-羟基扁桃酸、香草醛、地骨皮乙素等酚类物质;水杨酸、苯甲酸、邻乙酰氨基酚等苯环型化合物;萘类、蒽类和酚酯类物质),占比3.070%。表 1 代谢物分类及占比Table 1. Proportions of classified metabolites代谢物种类
Metabolite class数量
Number占比
Percentage/%脂质和类脂分子
Lipids and lipid-like molecules119 4.349 莽草酸和苯丙烷类
Shikimates and Phenylpropanoids112 4.094 苯丙烷和聚酮
Phenylpropanoids and polyketides96 3.509 有机杂环化合物
Organoheterocyclic compounds89 3.253 苯环型化合物
Benzenoids84 3.070 脂肪酸
Fatty acids68 2.485 有机酸及其衍生物
Organic acids and derivatives65 2.376 萜类化合物
Terpenoids65 2.376 生物碱类
Alkaloids59 2.156 有机含氧化合物
Organic oxygen compounds44 1.608 氨基酸和肽核苷
Amino acids and Peptides32 1.170 核苷酸和类似物
Nucleosides, nucleotides, and analogues29 1.060 碳水化合物
Carbohydrates19 0.694 聚酮
Polyketides16 0.585 生物碱及其衍生物
Alkaloids and derivatives15 0.548 有机氮化合物
Organic nitrogen compounds9 0.329 木脂素、新木脂素及相关化合物
Lignans, neolignans and related compounds5 0.183 其他 Others 1810 66.155 共计 Total 2736 100.000 2.3 N-25发酵前后差异代谢物筛选
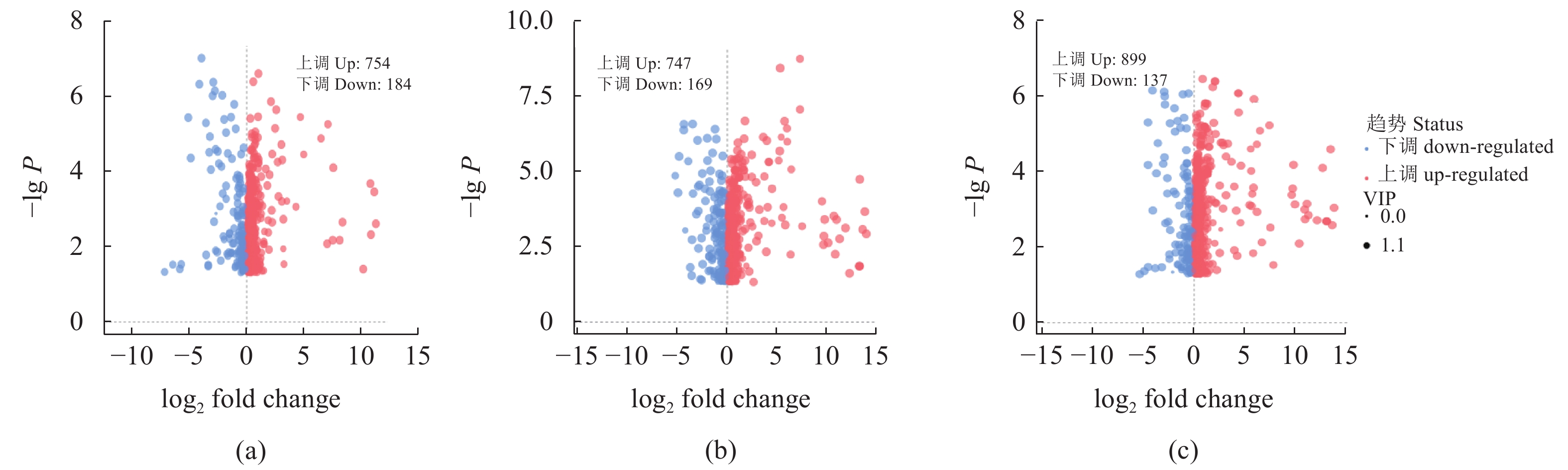
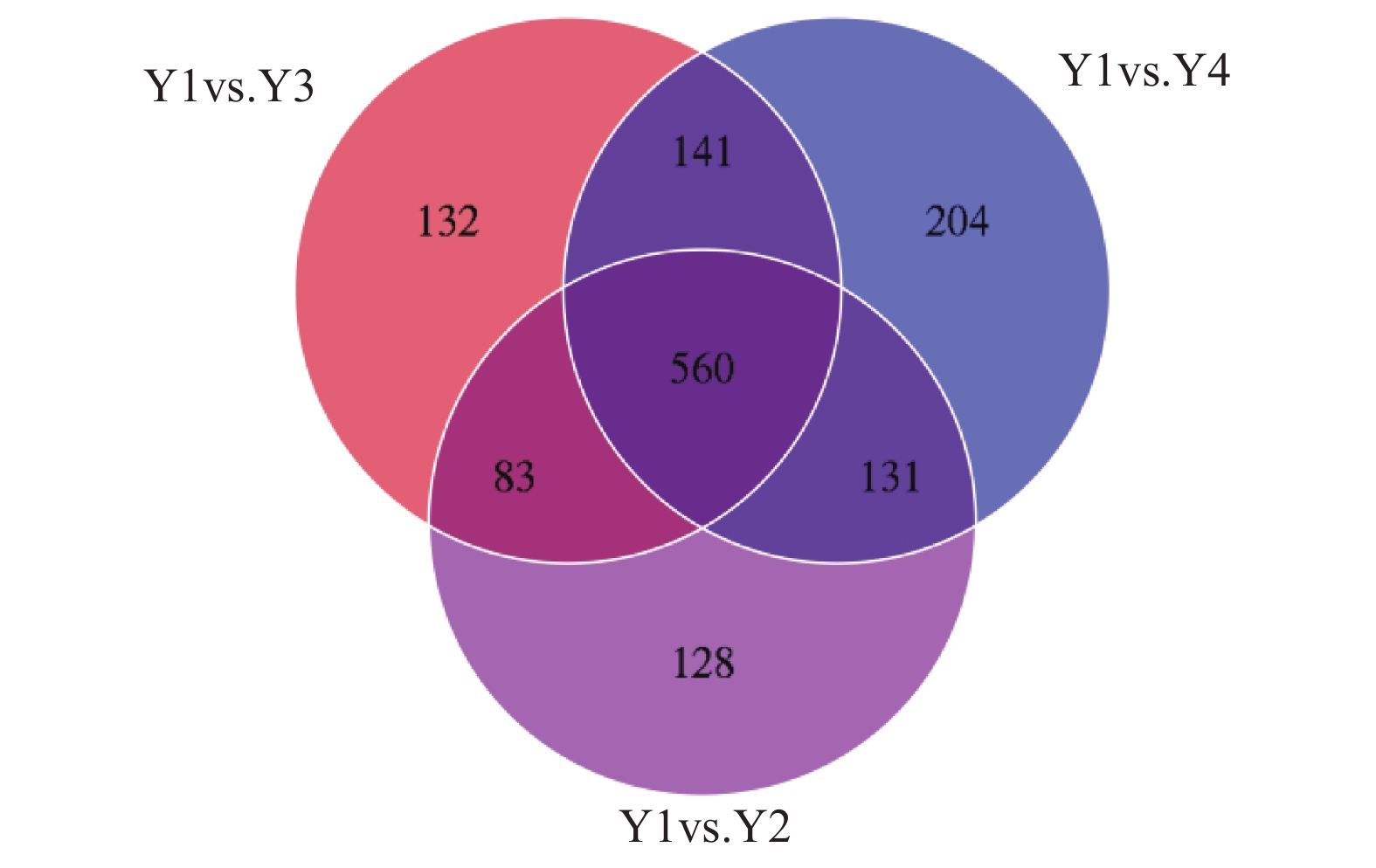
根据OPLS-DA结果,以VIP>1且P<0.05作为筛选条件,如图3所示,Y2比Y1组得到938个标志性差异代谢物,其中上调的有754个,下调的有184个;Y3比Y1组得到916个标志性差异代谢物,其中上调的有747个,下调的有169个;Y4比Y1组得到1036个标志性差异代谢物,其中上调的有899个,下调的有137个。图4展示了各组间差异代谢物之间的关系,三组间共同的差异代谢物有560个,Y2组特有差异代谢物为128种,包括3-氰基-L-丙氨酸、丙氨酸、2-萘酚、甘氨酸、甘露三糖、10-羟基癸酸等,Y3组有132种特有差异代谢物,包括康柠酸、月桂酸、3-羟基-3-甲基-2-氧代戊酸酯、异苹果酸、2-乙酰乳酸、L-亮氨酸等;Y4组特有的差异代谢物有204种,包括2-异丙基苹果酸、3-异丙基苹果酸、丙酮酸、L-异亮氨酸、L-缬氨酸等。
![]() 图 3 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的差异代谢物筛选火山图(a)Y1 vs.Y2;(b)Y1vs.Y3;(c)Y1vs.Y4。横坐标代表该组对比各物质的倍数变化(取以2为底的对数),纵坐标表示 t 检验的 P值(取以 10 为底对数的负数)。Figure 3. Volcanic diagram of differential metabolite screening between Y1 and Y2, Y1 and Y3, and Y1 and Y4(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4; horizontal axis: comparison of objects in same group on multiple quality changes (value in log2); vertical axis: P-values in student t-test (negative value of log10).
图 3 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的差异代谢物筛选火山图(a)Y1 vs.Y2;(b)Y1vs.Y3;(c)Y1vs.Y4。横坐标代表该组对比各物质的倍数变化(取以2为底的对数),纵坐标表示 t 检验的 P值(取以 10 为底对数的负数)。Figure 3. Volcanic diagram of differential metabolite screening between Y1 and Y2, Y1 and Y3, and Y1 and Y4(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4; horizontal axis: comparison of objects in same group on multiple quality changes (value in log2); vertical axis: P-values in student t-test (negative value of log10).![]() 图 4 Y1 vs.Y2、Y1 vs.Y3、Y1 vs.Y4的维恩图Figure 4. Venn diagram of Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4
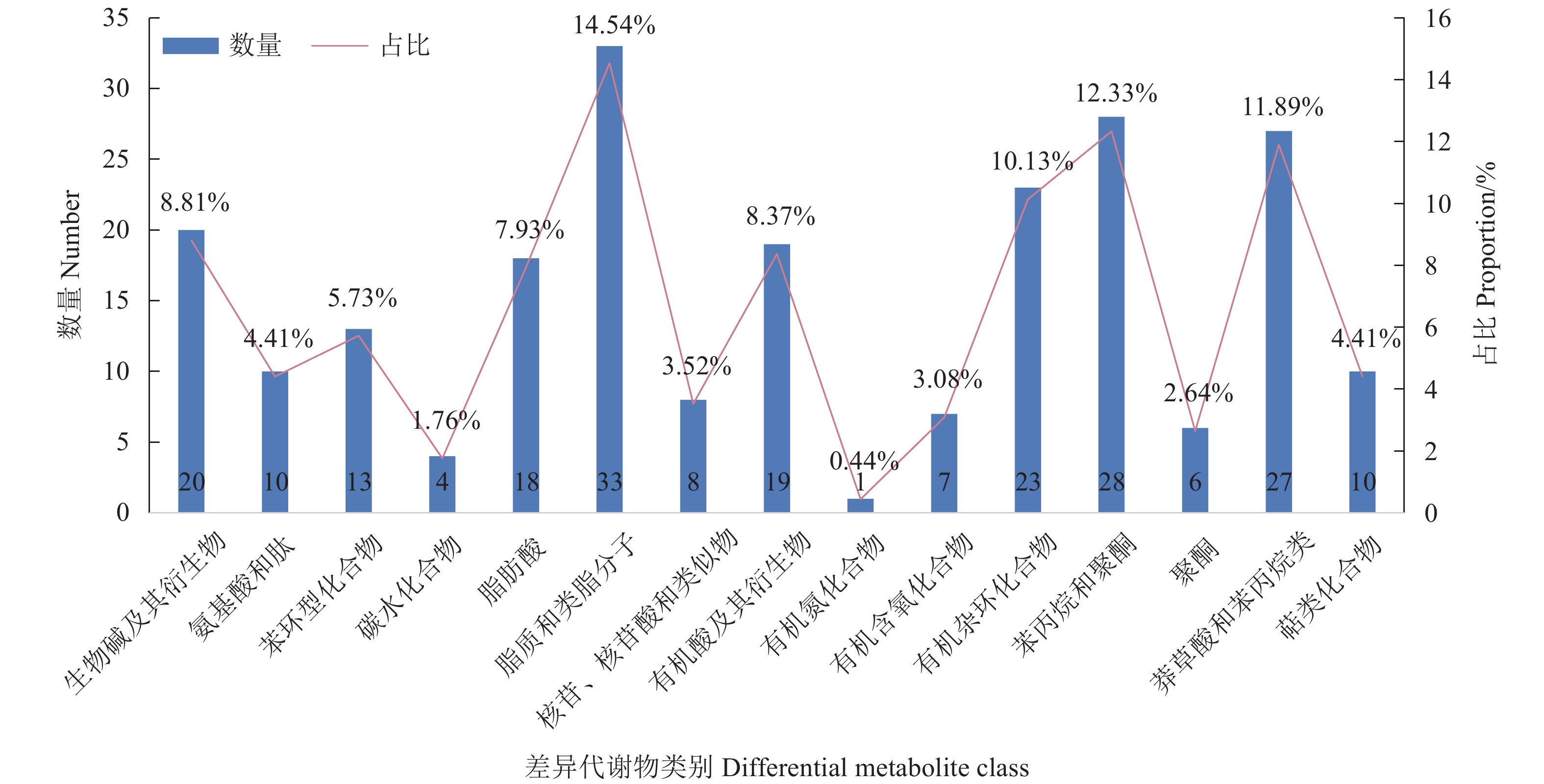
图 4 Y1 vs.Y2、Y1 vs.Y3、Y1 vs.Y4的维恩图Figure 4. Venn diagram of Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4为探究N-25对党参渣成分的影响,试验选取3个组共有的差异代谢物进行分析,其中被注释的差异代谢物有227个,图5展示了主要差异代谢物的分类与占比,可观察到主要差异代谢物有脂质和类脂分子、苯丙素烷和聚酮化合物、莽草酸和苯丙烷类、有机杂环化合物、生物碱及其衍生物、有机酸及衍生物以及脂肪酸。为了进一步观察植物乳杆菌N-25发酵引起的活性成分的变化,分别制作黄酮类化合物、木脂素、生物碱及其衍生物、有机酸及其衍生物的热图(图6)。
![]() 图 5 Y1 vs. Y2,Y1 vs. Y3,Y1 vs. Y4共有差异代谢物分类占比图Figure 5. Proportions of classes of differential metabolite shared by Y1 and Y2, Y1 and Y3, and Y1 and Y4
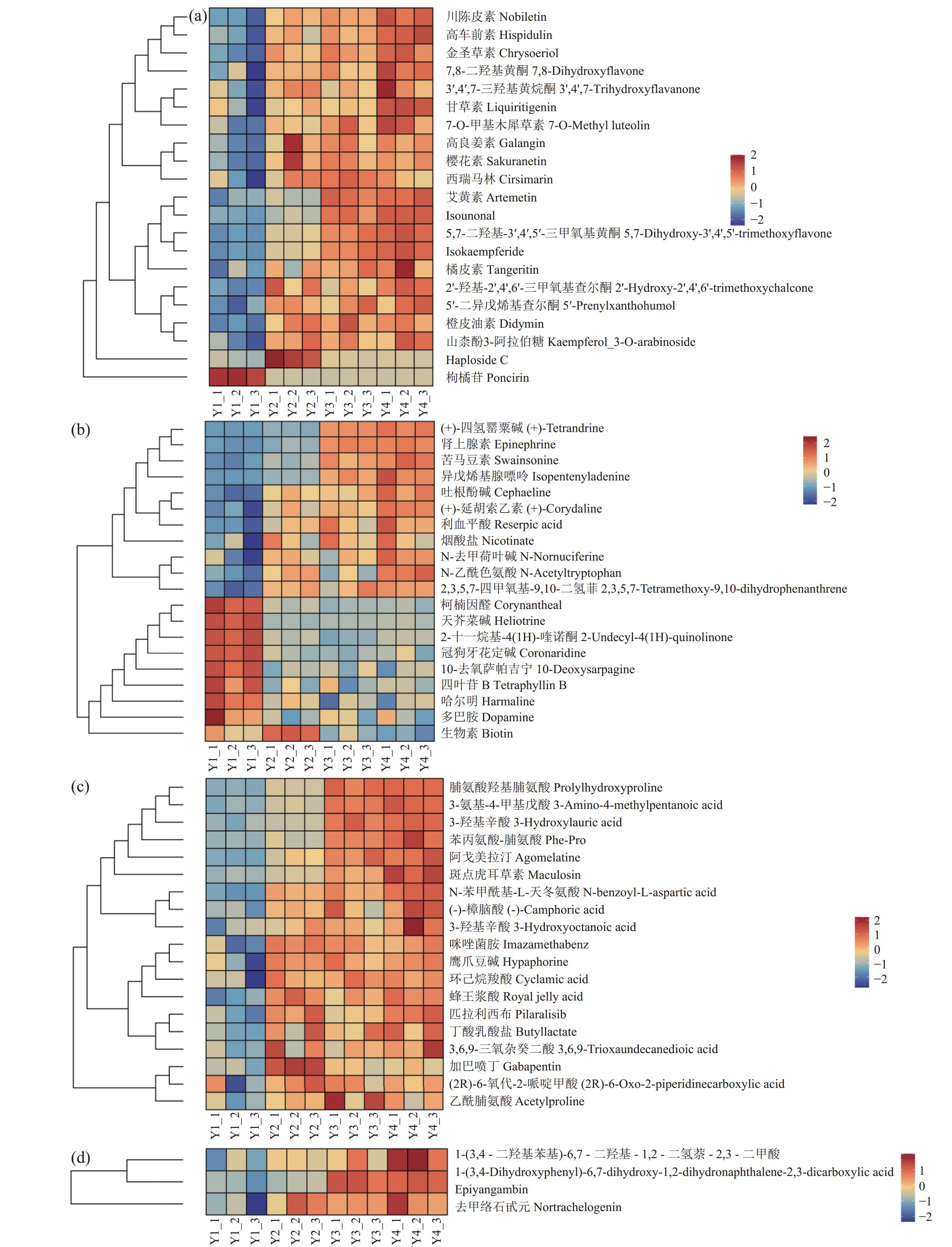
图 5 Y1 vs. Y2,Y1 vs. Y3,Y1 vs. Y4共有差异代谢物分类占比图Figure 5. Proportions of classes of differential metabolite shared by Y1 and Y2, Y1 and Y3, and Y1 and Y4![]() 图 6 Y1vs.Y2、Y1vs.Y3、Y1vs.Y4主要差异代谢物成分热图(a)黄酮化合物;(b)生物碱及衍生物;(c)有机酸及其衍生物;(d)木脂素。Figure 6. Heatmap of major differential metabolites in Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4(a) flavonoids; (b) alkaloids and derivatives; (c): organic acids and derivatives; (d) lignan.
图 6 Y1vs.Y2、Y1vs.Y3、Y1vs.Y4主要差异代谢物成分热图(a)黄酮化合物;(b)生物碱及衍生物;(c)有机酸及其衍生物;(d)木脂素。Figure 6. Heatmap of major differential metabolites in Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4(a) flavonoids; (b) alkaloids and derivatives; (c): organic acids and derivatives; (d) lignan.如图6所示,经N-25发酵后(图6),鉴定到川陈皮素、7-O-甲基木樨草素、高车前素、金圣草素和7,8-二羟基黄酮等18种黄酮类化合物含量均增加,较对照含量增加1.13倍;注释到的生物碱共20种,其中苦马豆素、烟酸盐、肾上腺素、(+)-四氢罂粟碱、异戊烯基腺嘌呤、吐根酚碱、(+)-延胡索乙素、利血平酸含量较对照增加1.01倍;鉴定到的3-羟基月桂酸、蜂王浆酸和(-)-樟脑酸等19种有机酸及其衍生物的相对含量较对照组增加1.16倍;另外,Epiyangambin、去甲络石甙元和1-(3,4-二羟基苯基)-6,7-二羟基-1,2-二氢萘-2,3-二甲酸3种木脂素在发酵后的党参渣中含量较对照增加1.04倍。
2.4 KEGG代谢通路分析
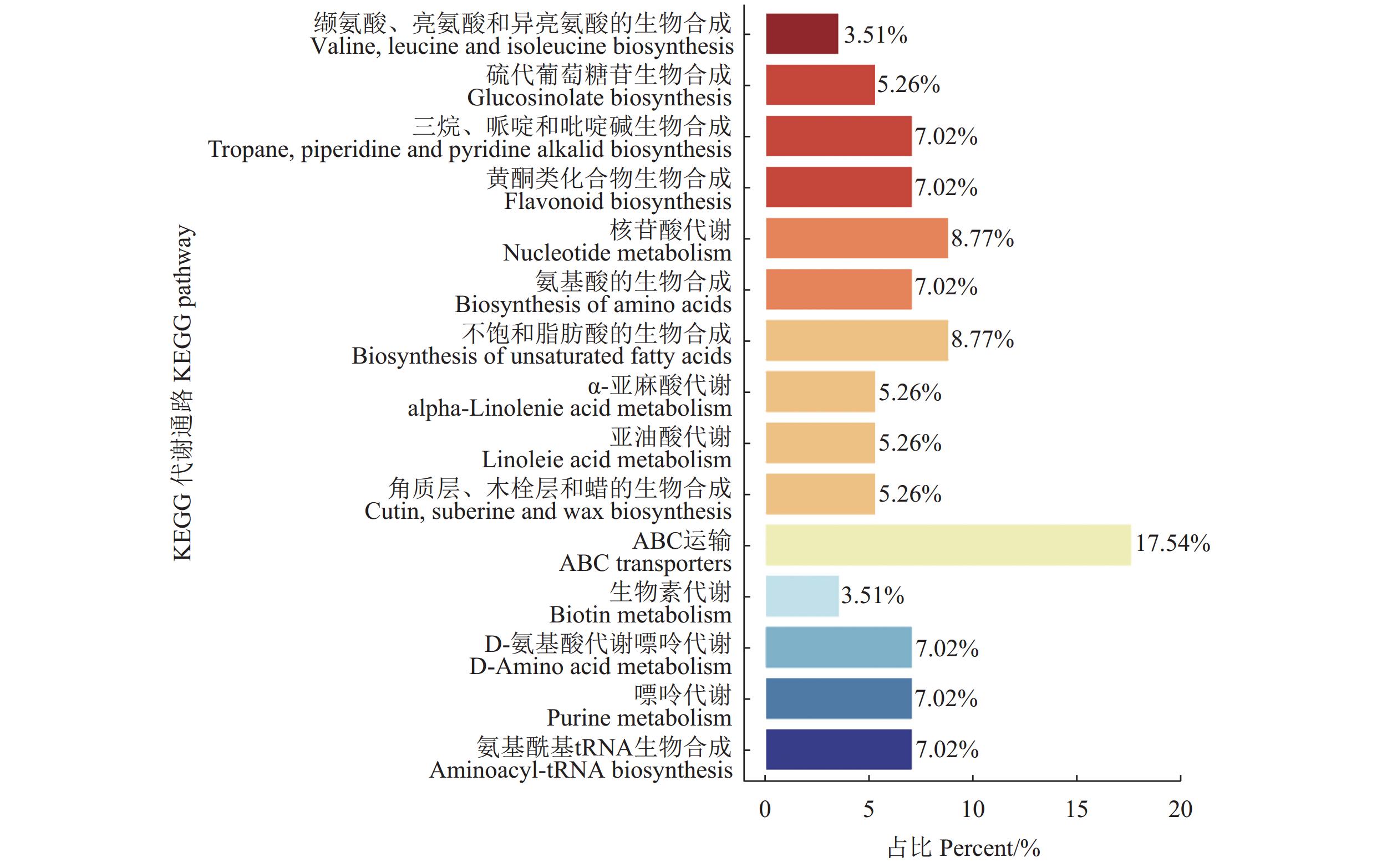
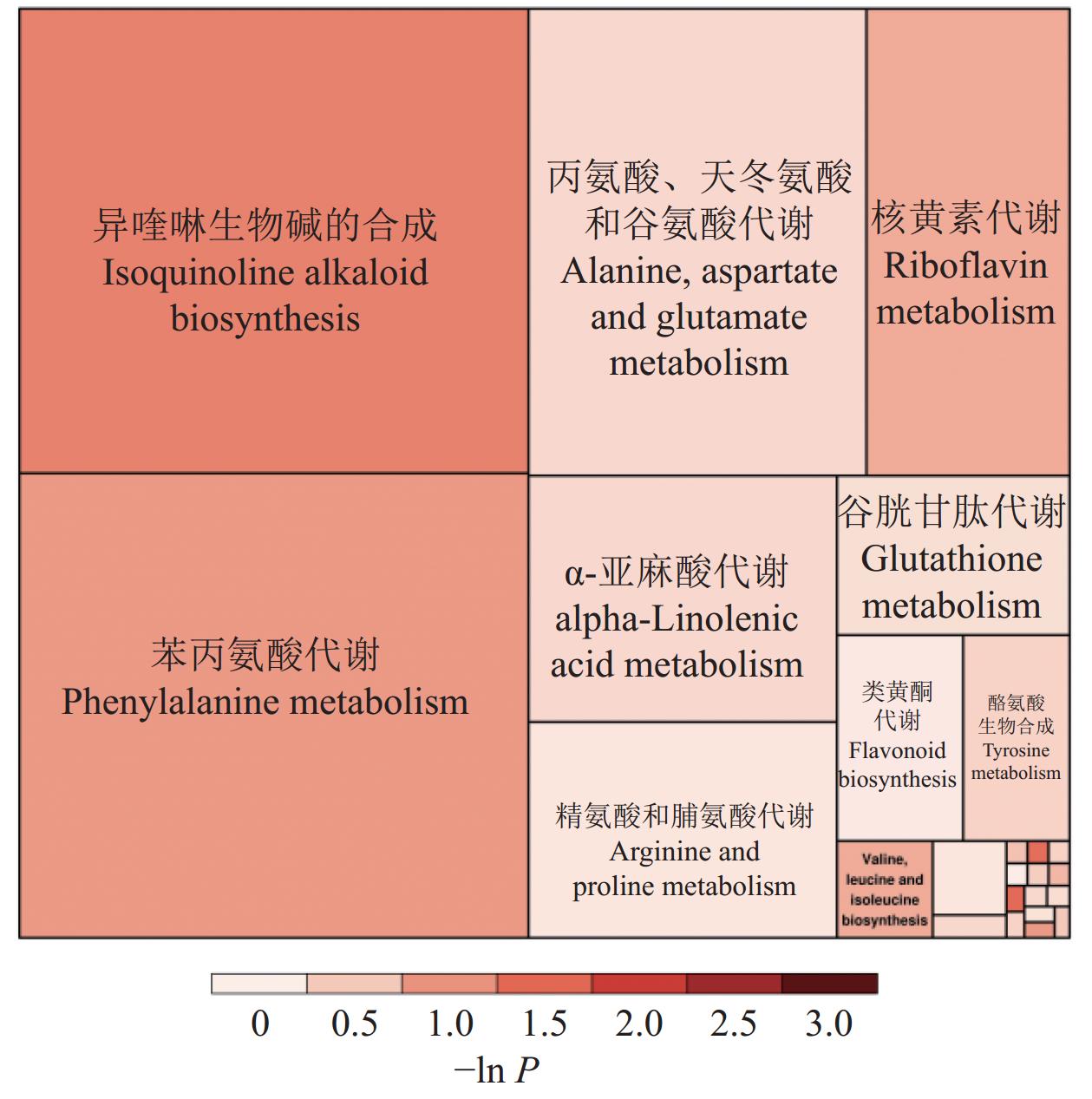
将筛选到的三组间共同的差异代谢物与KEGG数据库比对并进行通路富集分析。结果表明,在筛选出的560种具有显著差异代谢物中被KEGG注释到的共有56条通路。图7展示了差异代谢物富集到的部分通路,其中20.33%差异代谢物富集在代谢通路,13.19%差异代谢物富集在次级代谢物的生成通路,5.5%富集在ABC运输通路,3.3%富集在辅因子的生物合成通路,2.7%富集在核苷酸代谢,2.7%富集在不饱和脂肪酸的生成通路。其中关键通路包括异喹啉生物碱生物合成通路,苯丙氨酸代谢通路,核黄素代谢,丙氨酸、天冬氨酸和谷氨酸代谢通路,α-亚麻酸代谢通路(图8)。
![]() 图 7 Y1 vs.Y2、 Y1 vs.Y3、 Y1 vs.Y4组间共同差异代谢物KEGG分类图Figure 7. KEGG classification of common differential metabolites between Y1 and Y2, Y1 and Y3, and Y1 and Y4
图 7 Y1 vs.Y2、 Y1 vs.Y3、 Y1 vs.Y4组间共同差异代谢物KEGG分类图Figure 7. KEGG classification of common differential metabolites between Y1 and Y2, Y1 and Y3, and Y1 and Y4![]() 图 8 Y1 vs.Y2、 Y1 vs.Y3、 Y1 vs.Y4组间共同差异代谢物通路分析Figure 8. Common differential metabolite pathways between Y1 and Y2, Y1 and Y3, and Y1 and Y4
图 8 Y1 vs.Y2、 Y1 vs.Y3、 Y1 vs.Y4组间共同差异代谢物通路分析Figure 8. Common differential metabolite pathways between Y1 and Y2, Y1 and Y3, and Y1 and Y42.5 特有差异代谢物KEGG通路分析
如图9所示,1%N-25发酵党参渣组,其代谢物富集在ABC运输、丁酸盐代谢、烟酸和烟酰胺代谢通路上;3%N-25发酵党参组,其代谢物富集在C5受体二元酸代谢、缬氨酸、亮氨酸、异亮氨酸生物合成、丙氨酸、天冬氨酸和谷氨酸代谢通路上;5%N-25发酵党参组,其代谢物富集在次生代谢物的生物合成、咖啡因代谢、缬氨酸、亮氨酸、异亮氨酸生物合成代谢通路上。
![]() 图 9 Y1 vs.Y2、Y1 vs.Y3、Y1 vs.Y4组间特有差异代谢物KEGG分类(a)Y1vs.Y2,(b)Y1vs.Y3,(c)Y1vs.Y4。Figure 9. KEGG classification of unique metabolites between Y1 and Y2, Y1 and Y2, and Y1 and Y2(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.
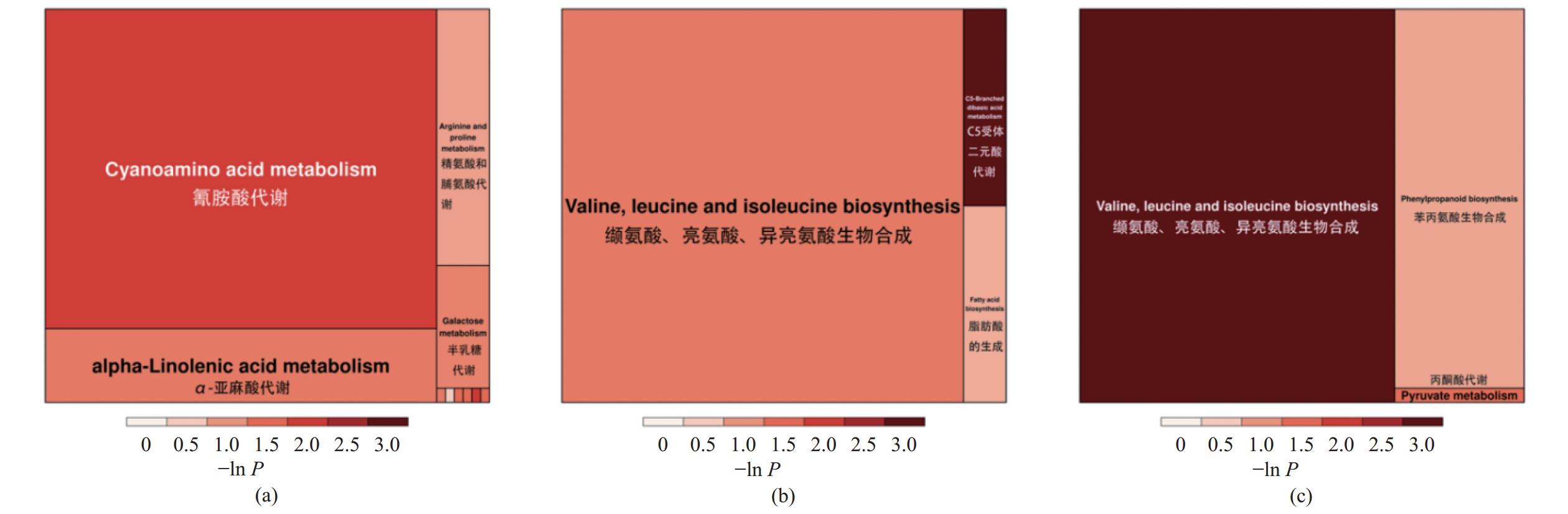
图 9 Y1 vs.Y2、Y1 vs.Y3、Y1 vs.Y4组间特有差异代谢物KEGG分类(a)Y1vs.Y2,(b)Y1vs.Y3,(c)Y1vs.Y4。Figure 9. KEGG classification of unique metabolites between Y1 and Y2, Y1 and Y2, and Y1 and Y2(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.为进一步研究不同接菌量的N-25发酵对党参渣代谢物及其相关代谢通路的影响,通过对差异代谢物所在通路的综合分析(包括富集分析和拓扑分析),对通路进行进一步的筛选,三组差异代谢物相关性最高的关键通路如图10所示,Y1 vs.Y2特异性差异代谢物集中的关键通路主要为氰氨酸代谢通路,Y1 vs. Y3特异性差异代谢物集中的关键通路主要为缬氨酸、亮氨酸、异亮氨酸生物合成,Y1 vs. Y4特异性差异代谢物集中的关键通路主要为缬氨酸、亮氨酸、异亮氨酸生物合成。
![]() 图 10 Y1 vs. Y2、Y1 vs. Y3、Y1 vs. Y4组间特有差异代谢物通路分析(a)Y1 vs.Y2,(b)Y1 vs.Y3,(c)Y1 vs.Y4。Figure 10. Metabolite pathways of unique metabolites between Y1 and Y2, Y1 and Y3, and Y1 and Y4(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.
图 10 Y1 vs. Y2、Y1 vs. Y3、Y1 vs. Y4组间特有差异代谢物通路分析(a)Y1 vs.Y2,(b)Y1 vs.Y3,(c)Y1 vs.Y4。Figure 10. Metabolite pathways of unique metabolites between Y1 and Y2, Y1 and Y3, and Y1 and Y4(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.3. 讨论与结论
本研究以N-25为发酵菌株,对党参渣进行单菌发酵,基于UHPLC-QE-MS的非靶代谢组学对发酵后的党参渣进行分析,结果显示,经过N-25发酵后,发酵产物共鉴定出
2737 种代谢物,包括32种羧酸及其衍生物,44种有机含氧化合物,55种脂肪酰基,70种黄酮类物质,74种生物碱,65种萜类,68种脂肪酸,32种氨基酸等。Zeng等[23]在废弃党参中共鉴定出1508 个代谢物,463种被鉴定为DEMs,可分为42种羧酸及其衍生物、34种类固醇和衍生物甾体、33种有机含氧化合物、32种脂肪酰基、21种黄酮类和其他类、26种生物碱、12种萜类和其他化合物。相较前人研究,发酵产物中活性成分的种类和含量都有增加,表明N-25发酵能提高党参渣的活性物质的释放,对党参渣的资源化利用提供了一种可能的有效途径。贾旭森[24]利用酵母菌发酵党参渣,发现黄酮、三萜、氨基酸含量等均有不同程度的增加。本研究采用N-25发酵党参渣,其黄酮、三萜、氨基酸含量显著增加,且与前人相比,增加幅度更明显,表明N-25对党参渣的发酵展现出了更优异的效果。本研究中含量增加的黄酮化合物,如川陈皮素和木樨草素具有良好的抗炎抗肿瘤作用[25−27],提示黄酮化合物的积累可能与N-25发酵党参渣中抗炎效应存在潜在关联,但还需进一步验证。苦马豆素作为一种生物碱,具有抗病毒和抗肿瘤的作用[28];烟酸盐具有抗炎作用[29];Epiyangambin具有显著的降压作用,并在预防和治疗血栓形成方面具有潜在应用价值[30−31];去甲络石甙元有抗氧化、抗炎和抗癌作用[32−33];3-羟基月桂酸和蜂王浆酸已被证实具有抗炎活性[34−36],(-)-樟脑酸可显著抑制前列腺素E2(PGE2)、白三烯B4(LTB4)及IL-6等炎症介质的生物合成[37]。本研究中,以上活性物质的含量均明显增加,进一步表明N25发酵能有效提升党参渣中活性成分的相对含量。
本研究中N-25发酵党参渣涉及众多通路,包括异喹啉生物碱合成通路、苯丙氨酸代谢通路、α-亚麻酸代谢通路等,其中相关性最强的是异喹啉生物碱合成通路,发酵后党参渣中异喹啉生物碱合成相关代谢物(+)-延胡索乙素、(+)-粉防己碱、吐根酚碱含量明显增加,多巴胺含量明显下降,推测N-25发酵的作用下存在N-甲基转移酶催化作用和伯胺N-甲基转移酶(NCS)催化作用,酪氨酸脱羧酶(TYDC)脱羧作用,将多巴胺转变为不同的异喹啉生物碱[38]。因异喹啉生物碱具有良好的抗炎作用[39−40],故认为N-25发酵党参渣对异喹啉生物碱活性物质的生成有积极的作用。
本试验研究结果表明,不同剂量N-25可以诱导党参渣产生不同特定功能的代谢产物,并且这些产物所涉及的功能也有所不同。Y2组中关键的特有代谢物3-氰基-L-丙氨酸与关键通路氰氨酸代谢通路直接相关[41−42],起到对发酵产物的解毒作用。亮氨酸、异亮氨酸和缬氨酸是人体营养必需支链氨基酸,它们不仅是蛋白质的基本组成单位,而且在细胞信号和分子调节、糖脂代谢、凋亡和自噬等方面发挥着重要的生理作用[43];Y3组主要特有代谢物2-Methyl-maleate参与关键通路为亮氨酸、异亮氨酸和缬氨酸合成通路,且Y3组亮氨酸和缬氨酸含量显著上升;Y4组关键的特有代谢物3-异丙基苹果酸和2-异丙基苹果酸是其关键通路亮氨酸、异亮氨酸和缬氨酸合成通路途径中的重要中间体[44−45];Y4组中亮氨酸和缬氨酸含量高于Y3组。因此,不同的N-25接菌量会产生不同的活性物质,但其作用机制还需进一步挖掘。
综上,植物乳杆菌N-25发酵党参渣,促进了黄酮类化合物、木脂素、生物碱及其衍生物、有机酸及其衍生物等活性物质的生成与释放;其共有差异代谢物富集的显著性差异代谢通路为异喹啉生物碱生物合成。
-
![]()
图 1 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的PCA分析
(a)Y1 vs. Y2,(b)Y1 vs. Y3,(c)Y1 vs. Y4;图中横坐标t[1]P表示第一主成分的预测主成分得分,展示样本组间差异,纵坐标t[1]O表示正交主成分得分,展示样本组内差异。
Figure 1. PCA scores for Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4
(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4; horizontal axis t[1] P: predicted scores on 1st principal component showing differences between sample groups; vertical axis t[1] O: scores of orthogonal principal component showing differences within same sample group.
![]()
图 2 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的OPLS-DA模型得分
(a)Y1vs.Y2,(b)Y1vs.Y3,(c)Y1vs.Y4。
Figure 2. OPLS-DA model scores for Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4
(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.
![]()
图 3 Y1组对Y2组、Y1组对Y3组、Y1组对Y4组的差异代谢物筛选火山图
(a)Y1 vs.Y2;(b)Y1vs.Y3;(c)Y1vs.Y4。横坐标代表该组对比各物质的倍数变化(取以2为底的对数),纵坐标表示 t 检验的 P值(取以 10 为底对数的负数)。
Figure 3. Volcanic diagram of differential metabolite screening between Y1 and Y2, Y1 and Y3, and Y1 and Y4
(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4; horizontal axis: comparison of objects in same group on multiple quality changes (value in log2); vertical axis: P-values in student t-test (negative value of log10).
![]()
图 4 Y1 vs.Y2、Y1 vs.Y3、Y1 vs.Y4的维恩图
Figure 4. Venn diagram of Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4
![]()
图 5 Y1 vs. Y2,Y1 vs. Y3,Y1 vs. Y4共有差异代谢物分类占比图
Figure 5. Proportions of classes of differential metabolite shared by Y1 and Y2, Y1 and Y3, and Y1 and Y4
![]()
图 6 Y1vs.Y2、Y1vs.Y3、Y1vs.Y4主要差异代谢物成分热图
(a)黄酮化合物;(b)生物碱及衍生物;(c)有机酸及其衍生物;(d)木脂素。
Figure 6. Heatmap of major differential metabolites in Y1 vs. Y2, Y1 vs. Y3, and Y1 vs. Y4
(a) flavonoids; (b) alkaloids and derivatives; (c): organic acids and derivatives; (d) lignan.
![]()
图 7 Y1 vs.Y2、 Y1 vs.Y3、 Y1 vs.Y4组间共同差异代谢物KEGG分类图
Figure 7. KEGG classification of common differential metabolites between Y1 and Y2, Y1 and Y3, and Y1 and Y4
![]()
图 8 Y1 vs.Y2、 Y1 vs.Y3、 Y1 vs.Y4组间共同差异代谢物通路分析
Figure 8. Common differential metabolite pathways between Y1 and Y2, Y1 and Y3, and Y1 and Y4
![]()
图 9 Y1 vs.Y2、Y1 vs.Y3、Y1 vs.Y4组间特有差异代谢物KEGG分类
(a)Y1vs.Y2,(b)Y1vs.Y3,(c)Y1vs.Y4。
Figure 9. KEGG classification of unique metabolites between Y1 and Y2, Y1 and Y2, and Y1 and Y2
(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.
![]()
图 10 Y1 vs. Y2、Y1 vs. Y3、Y1 vs. Y4组间特有差异代谢物通路分析
(a)Y1 vs.Y2,(b)Y1 vs.Y3,(c)Y1 vs.Y4。
Figure 10. Metabolite pathways of unique metabolites between Y1 and Y2, Y1 and Y3, and Y1 and Y4
(a) Y1 vs. Y2; (b) Y1 vs. Y3; (c) Y1 vs. Y4.
表 1 代谢物分类及占比
Table 1 Proportions of classified metabolites
代谢物种类
Metabolite class数量
Number占比
Percentage/%脂质和类脂分子
Lipids and lipid-like molecules119 4.349 莽草酸和苯丙烷类
Shikimates and Phenylpropanoids112 4.094 苯丙烷和聚酮
Phenylpropanoids and polyketides96 3.509 有机杂环化合物
Organoheterocyclic compounds89 3.253 苯环型化合物
Benzenoids84 3.070 脂肪酸
Fatty acids68 2.485 有机酸及其衍生物
Organic acids and derivatives65 2.376 萜类化合物
Terpenoids65 2.376 生物碱类
Alkaloids59 2.156 有机含氧化合物
Organic oxygen compounds44 1.608 氨基酸和肽核苷
Amino acids and Peptides32 1.170 核苷酸和类似物
Nucleosides, nucleotides, and analogues29 1.060 碳水化合物
Carbohydrates19 0.694 聚酮
Polyketides16 0.585 生物碱及其衍生物
Alkaloids and derivatives15 0.548 有机氮化合物
Organic nitrogen compounds9 0.329 木脂素、新木脂素及相关化合物
Lignans, neolignans and related compounds5 0.183 其他 Others 1810 66.155 共计 Total 2736 100.000  下载: 导出CSV
下载: 导出CSV
-
[1] 冯亚静,王晓霞,庄鹏宇,等. 党参的化学成分研究[J]. 中国中药杂志,2017,42(1) :135−139. FENG Y J,WANG X X,ZHUANG P Y,t al. Study on chemical constituents of Codonopsis pilosula[J]. China Journal of Chinese Materia Medica,2017, 42(1) :135−139.(in Chinese)
[2] ENDO K,TAGUCHI T,TAGUCHI F,et al. Antiinflammatory principles of Atractylodes rhizomes[J]. Chemical & Pharmaceutical Bulletin,1979, 27(12) :2954−2958.
[3] 2022—2026年党参市场调查研究报告[M]. 北京:中研普华研究院,2022. Market Research Report on Codonopsis pilosula from 2022 to 2026 [M]. Beijing:Zhongyan Puhua Research Institute,2022.
[4] INOUE M,SUZUKI R,SAKAGUCHI N,et al. Selective induction of cell death in cancer cells by Gallic acid[J]. Biological & Pharmaceutical Bulletin,1995,18(11) :1526−1530.
[5] 解超平. 饲料中添加发酵中药渣对铁脚麻鸡肉质性状的改善[J]. 畜禽业,2019,30(4) :9−10,12 . XIE C P. Improvement of meat quality of iron-footed hemp chicken by adding fermented Chinese medicine residue to feed[J]. Livestock and Poultry Industry,2019, 30(4) :9−10,12.
[6] 张晓燕,龚苏晓,张铁军,等. 药用植物废弃物再利用研究现状[J]. 中草药,2016,47(7) :1225−1229. ZHANG X Y,GONG S X,ZHANG T J,et al. Research status of recycling medicinal plant waste[J]. Chinese Traditional and Herbal Drugs,2016,47(7) :1225−1229.(in Chinese)
[7] 吴金梅,胡迎利. 复方党参药对固态发酵的实验研究[J]. 现代牧业,2019,3(2) :6−8. DOI: 10.3969/j.issn.1008-3111.2019.02.002 WU J M,HU Y L. Experimental study on solid state fermentation by compound Codonopsis pilosula[J]. Modern Animal Husbandry,2019,3(2) :6−8.(in Chinese) DOI: 10.3969/j.issn.1008-3111.2019.02.002
[8] 罗志毅,黄新,包国荣. 大黄中蒽醌类成分清除氧自由基作用的研究[J]. 海峡药学,2009,21(12) :43−45. DOI: 10.3969/j.issn.1006-3765.2009.12.015 LUO Z Y,HUANG X,BAO G R. ESR Study on the scavenging effects of the free anthraquinones from Rheum palmatum L. on superoxide anion radical[J]. Strait Pharmaceutical Journal,2009,21(12) :43−45.(in Chinese) DOI: 10.3969/j.issn.1006-3765.2009.12.015
[9] LI H C,TAN P,LEI W Z,et al. Effect of microwave-puffed on Auricularia auricula polysaccharide and probiotic fermentation on its biotransformation and quality characteristics during storage period[J]. International Journal of Biological Macromolecules,2024,281(Pt 3) :136448.
[10] 孟晓燕. 发酵中药渣对犊牛生长发育和健康状况的影响[J]. 山东畜牧兽医,2024,45(9) :14−15,18. DOI: 10.3969/j.issn.1007-1733.2024.09.005 MENG X Y. Effect of residue in fermentation on growth and health status of calves[J]. Shandong Journal of Animal Science and Veterinary Medicine,2024,45(9) :14−15,18.(in Chinese) DOI: 10.3969/j.issn.1007-1733.2024.09.005
[11] 李华伟. 四种发酵中药渣对母猪繁殖性能和仔猪生长性能的影响[D]. 武汉:武汉轻工大学,2017. LI H W. Effects of four kinds of fermented Chinese medicine residues on reproductive performance of sows and growth performance of piglets[D]. Wuhan:Wuhan Polytechnic University,2017.(in Chinese)
[12] 张余,赵生月,王颖颖,等. 植物乳杆菌发酵添加太子参的培养基优化及抗氧化活性研究[J]. 现代食品,2024,30(9) :187−196. ZHANG Y,ZHAO S Y,WANG Y Y,et al. Optimization of Lactobacillus plantarum fermentation medium incorporated with the Pseudostellaria heterophylla and investigation of the antioxidant activity of the generated fermentation broth[J]. Modern Food,2024,30(9) :187−196.(in Chinese)
[13] 张亚芹,李家军,李豪,等. 植物乳杆菌发酵四君子散对肉鸡生长性能的影响[J]. 山西农业科学,2024,52(2) :97−102. ZHANG Y Q,LI J J,LI H,et al. Effects of sijunzi powder fermentated by Lactobacillus plantarum on growth performance of broilers[J]. Journal of Shanxi Agricultural Sciences,2024,52(2) :97−102.(in Chinese)
[14] 侯楠楠,谢全喜,王倩,等. 植物乳杆菌发酵复方中药对阿魏酸和甘草次酸溶出率的影响[J]. 广东饲料,2024,33(3) :43−46. DOI: 10.3969/j.issn.1005-8613.2024.03.014 HOU N N,XIE Q X,WANG Q,et al. Effect of Lactobacillus plantarum fermentation compound Chinese medicine on dissolution rate of ferulic acid and glycyrrhetinic acid[J]. Guangdong Feed,2024,33(3) :43−46.(in Chinese) DOI: 10.3969/j.issn.1005-8613.2024.03.014
[15] 厉桓. 植物乳杆菌固态发酵黄芪互作研究及在肉鸡上的应用[D]. 郑州:河南农业大学,2023. LI H. Study on the interaction of Astragalus membranaceus by solid-state fermentation of Lactobacillus plantarum and its application in broilers[D]. Zhengzhou:Henan Agricultural University,2023.(in Chinese)
[16] WANG J Y,ZHANG J,GUO H N,et al. Optimization of exopolysaccharide produced by Lactobacillus plantarum R301 and its antioxidant and anti-inflammatory activities[J]. Foods,2023,12(13) :2481. DOI: 10.3390/foods12132481
[17] FU H,ZHANG Y Z,AN Q,et al. Anti-photoaging effect of Rhodiola rosea fermented by Lactobacillus plantarum on UVA-damaged fibroblasts[J]. Nutrients,2022,14(11) :2324. DOI: 10.3390/nu14112324
[18] EWEYS A S,ZHAO Y S,DARWESH O M. Improving the antioxidant and anticancer potential of Cinnamomum cassia via fermentation with Lactobacillus plantarum[J]. Biotechnology Reports,2022,36:e00768. DOI: 10.1016/j.btre.2022.e00768
[19] DONG J J,NA Y X,HOU A J,et al. A review of the botany,ethnopharmacology,phytochemistry,analysis method and quality control,processing methods,pharmacological effects,pharmacokinetics and toxicity of Codonopsis Radix[J]. Frontiers in Pharmacology,2023,14:1162036. DOI: 10.3389/fphar.2023.1162036
[20] 林洁鑫,王鹏杰,金珊,等. 基于广泛靶向代谢组学的不同产地红茶代谢产物比较分析[J]. 食品工业科技,2022,43(2) :9−19. LIN J X,WANG P J,JIN S,et al. Comparative analysis of black tea metabolites from different origins based on extensively targeted metabolomics[J]. Science and Technology of Food Industry,2022,43(2) :9−19.(in Chinese)
[21] LI Z L,GUO Q,LIN F Q,et al. Lactobacillus plantarum supernatant inhibits growth of Riemerella anatipestifer and mediates intestinal antimicrobial defense in Muscovy ducks[J]. Poultry Science,2024,103(2) :103216. DOI: 10.1016/j.psj.2023.103216
[22] 秦楠,张娜郡,陈超,等. 党参发酵工艺及成分变化研究[J]. 中华中医药学刊,2023,41(6) :232–236,277–278. QIN N,ZHANG N J,CHEN C,et al. Study on fermentation process and component change of Dangshen(Codonopsis pilosula) [J]. Chinese Archives of Traditional Chinese Medicine,2023,41(6) :232–236,277–278.(in Chinese)
[23] ZENG X,LI J X,LYU X K,et al. Untargeted metabolomics reveals multiple phytometabolites in the agricultural waste materials and medicinal materials of Codonopsis pilosula[J]. Frontiers in Plant Science,2022,12:814011. DOI: 10.3389/fpls.2021.814011
[24] 贾旭森. 新鲜白条党参酵母菌固体发酵工艺及其成分和抗氧化活性研究[D]. 兰州:兰州大学,2021. JIA X S. Study on solid fermentation technology,composition and antioxidant activity of fresh Codonopsis pilosula yeast[D]. Lanzhou:Lanzhou University,2021.(in Chinese)
[25] PANG Y S,ZHANG L,ZHONG Z T,et al. Nobiletin restores HFD-induced enteric nerve injury by regulating enteric glial activation and the GDNF/AKT/FOXO3a/P21 pathway[J]. Molecular Medicine,2024,30(1) :113. DOI: 10.1186/s10020-024-00841-8
[26] CAI S J,GOU Y D,CHEN Y Y,et al. Luteolin exerts anti-tumour immunity in hepatocellular carcinoma by accelerating CD8+ T lymphocyte infiltration[J]. Journal of Cellular and Molecular Medicine,2024,28(17) :e18535. DOI: 10.1111/jcmm.18535
[27] SU Y Y,FU X,ZHUANG P W. Untargeted metabolomics analysis of lactic acid bacteria fermented Acanthopanax senticosus with regard to regulated gut microbiota in mice[J]. Molecules,2024,29(17) :4074. DOI: 10.3390/molecules29174074
[28] ELBEIN A D,MOLYNEUX R J. Alkaloid glycosidase inhibitors[M]. Comprehensive Natural Products II. Amsterdam:Elsevier,2010:225–260.
[29] VARUN B V,VAITHEGI K,YI S,et al. Nature-inspired remodeling of (aza) indoles to meta-aminoaryl nicotinates for late-stage conjugation of vitamin B3 to (hetero) arylamines[J]. Nature Communications,2020,11(1) :6308. DOI: 10.1038/s41467-020-19610-2
[30] ARAÚJO I G A,SILVA D F,DO CARMO DE ALUSTAU M,et al. Calcium influx inhibition is involved in the hypotensive and vasorelaxant effects induced by yangambin[J]. Molecules,2014,19(5) :6863−6876. DOI: 10.3390/molecules19056863
[31] CASTRO-FARIA-NETO H C,MARTINS M A,SILVA P M,et al. Pharmacological profile of epiyangambin:A furofuran lignan with PAF antagonist activity[J]. Journal of Lipid Mediators,1993,7(1) :1−9.
[32] WANG H F,HUANG Z H,ZOU W,et al. Tracheloside,the main constituent of the total lignan extract from Trachelospermi Caulis,inhibited rheumatoid arthritis via IL-17/MAPK signaling pathway[J]. Fitoterapia,2025,180:106311. DOI: 10.1016/j.fitote.2024.106311
[33] PEUHU E,PAUL P,REMES M,et al. The antitumor lignan Nortrachelogenin sensitizes prostate cancer cells to TRAIL-induced cell death by inhibition of the Akt pathway and growth factor signaling[J]. Biochemical Pharmacology,2013,86(5) :571−583. DOI: 10.1016/j.bcp.2013.05.026
[34] IZQUIERDO-VEGA J,ARTEAGA-BADILLO D,SÁNCHEZ-GUTIÉRREZ M,et al. Organic acids from Roselle (Hibiscus sabdariffa L.) :A brief review of its pharmacological effects[J]. Biomedicines,2020,8(5) :100. DOI: 10.3390/biomedicines8050100
[35] KRAUSE D,SUH H S,TARASSISHIN L,et al. The tryptophan metabolite 3-hydroxyanthranilic acid plays anti-inflammatory and neuroprotective roles during inflammation role of hemeoxygenase-1[J]. The American Journal of Pathology,2011,179(3) :1360−1372. DOI: 10.1016/j.ajpath.2011.05.048
[36] SUGIYAMA T,TAKAHASHI K,MORI H. Royal jelly acid,10-hydroxy-trans-2-decenoic acid,as a modulator of the innate immune responses[J]. Endocrine,Metabolic & Immune Disorders Drug Targets,2012,12(4) :368–376.
[37] 殷典贺,王秋月. 熊果酸抗炎、抗氧化作用及其机制的研究进展[J]. 国际呼吸杂志,2015(13) :1022−1025. DOI: 10.3760/cma.j.issn.1673-436X.2015.13.015 YIN D H,WANG Q Y. Research progress on anti-inflammatory,antioxidant effects and its mechanism of ursolic acid[J]. International Journal of Respiration,2015(13) :1022−1025.(in Chinese) DOI: 10.3760/cma.j.issn.1673-436X.2015.13.015
[38] 赵艳妹,刘林娅,鲁明秋,等. 异黄酮生物合成通路及关键酶研究进展[J]. 食品与发酵工业,2024,50(2) :343−353. ZHAO Y M,LIU L Y,LU M Q,et al. Advances on pathway of isoflavone biosynthesis and relevant key enzymes[J]. Food and Fermentation Industries,2024,50(2) :343−353.(in Chinese)
[39] 李宗恒,张雪芳,陈延华,等. 基于16sRNA测序分析亚油酸对小鼠肠道菌群的影响[J]. 安徽医科大学学报,2024,59(7) :1116−1122. LI Z H,ZHANG X F,CHEN Y H,et al. The effects of linoleic acid on intestinal flora in mice were analyzed based on 16sRNA sequencing[J]. Acta Universitatis Medicinalis Anhui,2024,59(7) :1116−1122.(in Chinese)
[40] GAO W,GUAN P,GAO W P,et al. Lycorine attenuates lipopolysaccharide-induced inflammation and intestinal epithelial barrier dysfunction in Caco-2 cells through inhibiting the STING/NF-κB pathway[J]. Pakistan Journal of Pharmaceutical Sciences,2024,37(6) :1443−1454.(in Chinese)
[41] GUPTA N,BALOMAJUMDER C,AGARWAL V K. Enzymatic mechanism and biochemistry for cyanide degradation:A review[J]. Journal of Hazardous Materials,2010,176(1/2/3) :1−13.
[42] MACHINGURA M,SALOMON E,JEZ J M,et al. The β-cyanoalanine synthase pathway:Beyond cyanide detoxification[J]. Plant,Cell & Environment,2016,39(10) :2329–2341.
[43] PIOTROWSKI M,VOLMER J J. Cyanide metabolism in higher plants:Cyanoalanine hydratase is a NIT4 homolog[J]. Plant Molecular Biology,2006,61(1) :111−122.
[44] MIGGIANO R,MARTIGNON S,MINASSI A,et al. Crystal structure of Haemophilus influenzae 3-isopropylmalate dehydrogenase (LeuB) in complex with the inhibitor O-isobutenyl oxalylhydroxamate[J]. Biochemical and Biophysical Research Communications,2020,524(4) :996−1002. DOI: 10.1016/j.bbrc.2020.02.022
[45] BURKE G,FIEHN O,MORAN N. Effects of facultative symbionts and heat stress on the metabolome of pea aphids[J]. The ISME Journal,2010,4(2) :242−252. DOI: 10.1038/ismej.2009.114

计量
- 文章访问数: 32
- HTML全文浏览量: 3
- PDF下载量: 3
