

Cloning and Biological Characteristics of PpCaMK1 in Pseudocohnilembus persalinus
-
摘要:目的
钙调素依赖蛋白激酶(calmodulin-dependent protein kinases, CaMKs)是钙离子介导的信号转导通路中的关键组成部分,可能成为治疗盾纤虫病的药物靶点。本研究试图克隆水滴伪康纤虫(Pseudocohnilembus persalinus)的钙调素依赖蛋白激酶基因,分析其蛋白结构、功能,并基于该蛋白的氨基酸序列构建相关纤毛虫的系统进化树,以阐明水滴伪康纤虫CaMK的进化特征及与相关纤毛虫的系统发育关系。
方法参考水滴伪康纤虫RNA-seq测序结果,利用cDNA末端快速克隆(RACE)技术获得水滴伪康纤虫钙调素依赖蛋白激酶基因的全长cDNA序列,将其命名为PpCaMK1;利用生物信息学方法对该基因的序列特征、基因结构进行分析,预测该基因编码蛋白的理化性质、结构域及蛋白结构等,并构建其与相关纤毛虫CaMK的系统发育树。
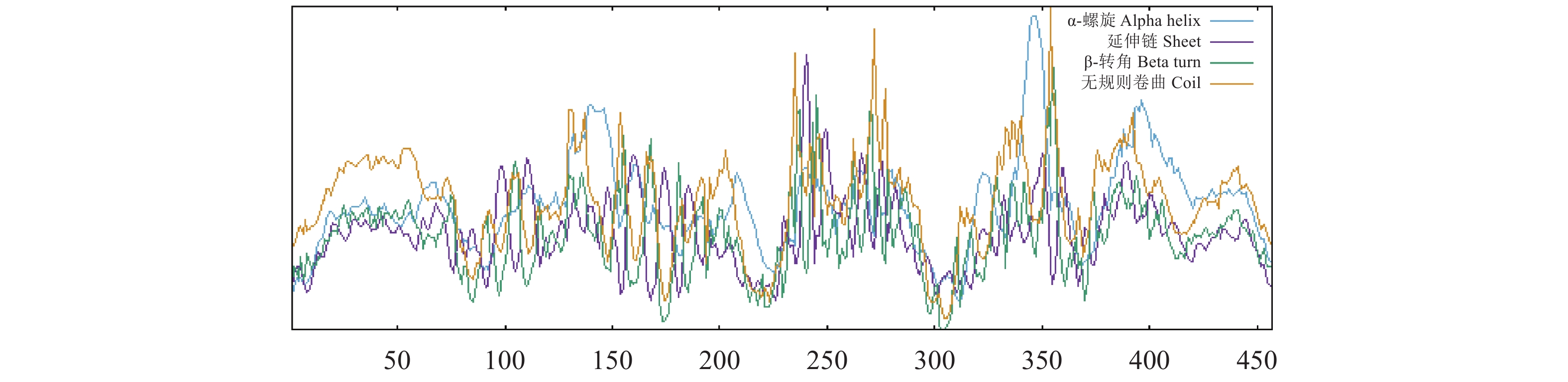
结果PpCaMK1基因包含有9个外显子和8个内含子,其cDNA全长为1 737 bp(GenBank序列号:PQ278249),其5′-UTR长度为56 bp,3′-UTR长度为307 bp,ORF总长度为1 374 bp,编码457个氨基酸残基。PpCaMK1总亲水性平均值为−0.802,不稳定指数为54.78,为亲水性不稳定蛋白。亚细胞定位预测分析表明PpCaMK1存在于细胞质-细胞核中。该蛋白无跨膜结构域,无信号肽。蛋白质的二级结构分析结果显示该蛋白含有的无规卷曲占比达50.33%。构建的系统发育树显示,水滴伪康纤虫PpCaMK1与寡膜纲纤毛虫同源蛋白具有较近的亲缘关系。
结论本研究克隆了水滴伪康纤虫PpCaMK1基因的全长cDNA,阐明该基因的结构,并初步分析PpCaMK1蛋白的基本理化性质及结构特征,结果可作为进一步研究PpCaMK1功能的基础,并为深入探究水滴伪康纤虫钙调素依赖蛋白激酶基因家族提供参考。
Abstract:ObjectiveGene associated with the calmodulin-dependent protein kinases (CaMKs) from Pseudocohnilembus persalinus was cloned to study its protein structure and function as well as genetic relation and evolutionary characteristics for the development of treatment for the parasitic scuticociliatosis in aquaculture species.
MethodBased on the RNA-seq of P. persalinus, the full-length cDNA sequence of PpCaMK1 was obtained using the Rapid Amplification of cDNA Ends (RACE) technique. Bioinformatic methods were employed to analyze the features, structure, physicochemical properties, domains and phylogenetic relationships with other ciliates of the gene.
ResultPpCaMK1 contained 9 exons and 8 introns. The full cDNA length was 1 737 bp (GenBank accession number: PQ278249) including a 5'-UTR of 56 bp, a 3'-UTR of 307 bp, and an ORF of 1 374 bp and encoded 457 amino acids. It was a hydrophilic and unstable protein with an average overall hydrophilicity of −0.802 and an instability index of 54.78. Distributed within the cytoplasm and nucleus, PpCaMK1 notably lacked the transmembrane domains and signal peptides and had a secondary structure composed of a significant proportion (i.e., 50.33%) of random coils. P. persalinus closely related to other ciliates in the class Oligohymenophorea.
ConclusionThe full-length cDNA of PpCaMK1 was successfully cloned and the basic biological and physicochemical characteristics studied paving the way for further research on the development of an effective treatment of the devastating disease on marine lives in aquacultural environment caused by the parasite P. persalinus.
-
Keywords:
- Pseudocohnilembus persalinus /
- CaMK /
- gene cloning /
- bioinformatics
-
0. 引言
【研究意义】木薯(Manihot esculenta Crantz)为大戟科木薯属直立灌木状多年生热带作物,起源于拉丁美洲[1],被广泛种植于热带和亚热带地区,是全球第六大粮食作物[2],为全球10亿多人提供了主要的食物来源[3]。木薯在种植过程中,面临着多种逆境胁迫,其中木薯花叶病(Cassava mosaic disease, CMD)是木薯最严重的病毒性疾病之一,可导致木薯高达95%的产量损失,是木薯生产的主要制约因素[4]。鉴于此,运用现代生物技术来发掘与逆境胁迫相关的基因,增强木薯的抗逆性和抗病性,显得尤其重要。【前人研究进展】研究人员通过全基因组关联研究(Genome-wide association studies, GWAS)和双亲连锁图谱发现CMD的抗性主要是由12号染色体上的一个主要基因座赋予的,该基因被称为CMD2基因座[5,6]。同时,Rabbi等研究发现CMD2基因座中的两个候选基因MePOD10(Manes.12G076200)和MePOD58(Manes.12G076300)与CMD的抗性紧密相关[7]。过氧化物酶是广泛存在于各种动物、植物和微生物体内的一类氧化酶[8]。其按照来源可分为3类,分别是Class Ⅰ、Class Ⅱ和Class Ⅲ[9]。第一类POD(Class Ⅰ POD),作为广泛存在于植物、真菌以及原核生物中的一种过氧化物酶,其主要功能是清除体内过多的过氧化氢,并在此过程中调节细胞的氧化还原稳态[10]。第二类POD(Class Ⅱ POD)主要存在于真菌中,并在木质素的降解过程中发挥着至关重要的作用[11]。第三类POD(Class Ⅲ POD)是植物中特有的分泌型过氧化物酶,在抵抗逆境胁迫的过程中发挥着重要作用[12]。在植物的生长及发育过程中,Class Ⅲ POD涉及了很多的生理生化反应,如参与生长素的分解代谢、细胞壁的合成代谢[13]、活性氮的代谢[14]、防御病原体[15]、木质素化和木栓形成[16]、活性氧(Reactive oxygen species, ROS)清除、伤口愈合和植物抗毒素合成等对生物和非生物胁迫的防御反应[17]。【本研究切入点】前人通过GWAS在CMD2基因座找到的MePOD10和MePOD58尚未在木薯中探究其功能,且有关木薯POD基因克隆和功能分析的研究相对较少。【拟解决的关键问题】本研究拟克隆MePOD10基因,利用生物信息学工具对其编码蛋白序列进行分析,并构建原核表达载体确定其蛋白表达条件,为深入探究MePOD10蛋白的功能奠定理论基础。
1. 材料与方法
1.1 试验材料
选用木薯品种SC124作为试验材料,其种植于海南大学儋州校区种植实验苗圃基地(19°50'N, 109°49'E),原核表达载体pET28a由实验室自行保存。植物总RNA提取试剂盒、异丙基-β-D-硫代吡喃半乳糖苷(IPTG)、DNA回收试剂盒购自天根生化科技有限公司、His标签蛋白纯化试剂盒购自上海碧云天生物技术股份有限公司、POD酶活测定试剂盒购自南京建成生物工程研究所、愈创木酚购自上海麦克林生化科技股份有限公司等。
1.2 木薯总RNA的提取和反转录
从SC124健康植株上剪取适量新鲜叶片,将其剪碎置于研钵内,加入少许液氮研磨成细腻粉状,经RNA提取试剂盒提取木薯总RNA后,再反转为cDNA,−80 ℃保存备用。
1.3 基因的克隆
在Phytozome网站(https://phytozome.jgi.doe.gov)查询MePOD10基因(Manes.12G076300)的CDS序列,用Primer 5.0软件设计特异性引物(表1),用cDNA作为模板扩增MePOD10基因CDS序列。经电泳跑胶验证PCR产物有目的条带后,切胶回收目的条带并将其构建于pEASY-Blunt3载体上,转化后挑选边缘清晰、大小适中且明亮的单菌落,摇菌并提取质粒送测序,将测序正确的单克隆菌液于−40 ℃保存备用。
表 1 供试引物序列Table 1. Primers applied引物名称
Primer name引物序列
Primer sequence引物用途
Primer usageMePOD10-F
MePOD10-R5'-GGAATTCATGGGAGGTTTTGGTTCTTT-3'
5'-GGATCCTCAACTATTGATCACTGCAC-3'基因克隆
Gene cloningMePOD10-pET28a-F
MePOD10-pET28a-R5'-ATGGGTCGCGGATCCATGGGAGGTTTTGGTTCTTT-3'
5'-GTCGACGGAGCTCGAATTCCAGGTCCTCCTCTGAGA
TCAGCTTCTGCTCCTCACTATTGATCACTGCAC-3'原核表达
Prokaryotic expression1.4 生物信息学分析
MePOD10基因编码的蛋白理化特性由ExPASy(http://web.expasy.org/protparam/)分析,MePOD10蛋白保守结构域利用NCBI(https://www.ncbi.nlm.nih.gov/)的CDD数据库分析。MePOD10蛋白的二级和三级结构利用Prabi(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html)和SWISS-MODEL(https://swissmodel.expasy.org/interactive)预测。通过NCBI数据库BLAST搜索MePOD10基因同源序列,并确定其开放阅读框(Open reading frame, ORF)。由MEGA7.0翻译MePOD10基因CDS序列,与其他物种进行同源比对分析后构建系统发育进化树。
1.5 原核表达载体的构建
以MePOD10-Blunt3质粒为模板,扩增MePOD10-pET28a片段,将扩增好的片段以同源重组方法构建于pET28a载体上,转化后挑选边缘清晰、大小适中且明亮的单菌落,摇菌并提取质粒送测序,将测序正确的单克隆菌液−40 ℃保存备用。
1.6 重组蛋白的诱导表达
将构建成功的MePOD10-pET28a质粒转入BL21(DE3)感受态细胞中,选取PCR检测无误的单克隆作为表达菌,将表达菌接种于LB培养基中,利用紫外分光光度计将表达菌液初始D600调至0.2~0.3,置于37 ℃摇床,表达菌液摇至D600为0.6~0.8,加入IPTG至终浓度为1 mmol·L−1并置于37 ℃摇床,200 r·min−1条件下诱导。取诱导6 h的重组蛋白为样品,经离心后取上清和沉淀加入上样缓冲液后充分涡旋,SDS-PAGE检测。
1.7 重组蛋白的Western blotting验证
经SDS-PAGE电泳后,进行转膜,转完的膜经1×TBS 漂洗后置于15 mL的脱脂牛奶中,室温下摇床轻晃1 h。每隔10 min用适量的1×TBS洗膜3次,加入Myc抗体室温孵育2 h,每隔10 min用适量的1×TBS洗膜3次,加入二抗室温孵育1 h,最后用A、B显影液进行显影成像。
1.8 重组蛋白的纯化和酶活测定
选用上述诱导条件对MePOD10重组蛋白进行大量诱导,将诱导后的菌液离心,用磷酸缓冲溶液重悬菌体,功率25%、冰上超声破碎10 min后离心得到的上清用试剂盒纯化。用POD酶活试剂盒测定MePOD10蛋白酶活性,pET28a空载蛋白作对照,重复3次。以不同浓度愈创木酚(0、10、20、30、40、50 mmol·L−1)为底物设置5组试验,以磷酸缓冲溶液作对照调零,在反应温度37 ℃、pH5.8条件下,取5 mL反应混合液(磷酸缓冲溶液3 mL、不同浓度的愈创木酚1 mL和1 mL 2%过氧化氢,摇匀),每组各加入1 mL纯化后的MePOD10蛋白,立即摇匀,并迅速倒入比色皿中,于470 nm波长下比色,1 min后记录各组吸光度值∆A,重复3次。按下式计算酶反应初始速度V0:
V0=ΔA⋅Vtε⋅Δt⋅Vs⋅c ∆A为反应初速度阶段内吸光度的变化;Vt为提取酶液总体积(mL);ε为愈创木酚的摩尔吸光系数(mol·L−1·cm−1);∆t为反应时间;Vs为测定时取用蛋白体积(mL);c为蛋白浓度(mg·mL−1)。使用GraphPad Prism 8.0拟合酶动力学曲线并进行单因素方差分析。
2. 结果与分析
2.1 MePOD10基因克隆及鉴定
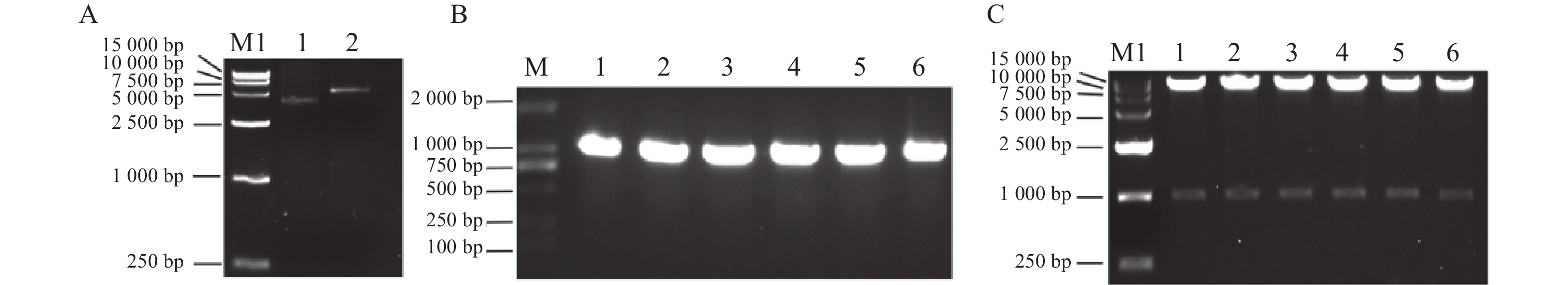
提取SC124木薯新鲜叶片总RNA并反转录成cDNA,以cDNA为模板进行PCR扩增,得到目的基因片段(图1A)。回收的目的片段构建于pEASY-Blunt3载体上,转入DH5α菌株中,菌落PCR选取阳性克隆(图1B),摇菌并提取质粒送测序。经序列比对显示,该阳性克隆长度为981 bp,编码326个氨基酸,鉴定为MePOD10基因。
![]() 图 1 MePOD10全长克隆M:DL2000 DNA标记;A:1~6 为MePOD10基因;B:1~6 为MePOD10基因阳性克隆。Figure 1. Cloned full-length MePOD10M: DL2000 DNA marker; A: 1-6: gene fragments of MePOD10; B: 1–6: positive clones after ligation transformation.
图 1 MePOD10全长克隆M:DL2000 DNA标记;A:1~6 为MePOD10基因;B:1~6 为MePOD10基因阳性克隆。Figure 1. Cloned full-length MePOD10M: DL2000 DNA marker; A: 1-6: gene fragments of MePOD10; B: 1–6: positive clones after ligation transformation.2.2 生物信息学分析
2.2.1 MePOD10编码蛋白理化性质和保守结构域分析
使用在线工具Expasy对MePOD10蛋白进行预测分析,结果显示MePOD10的分子式为C
1534 H2434 N434O481S13,分子量为35069.60 Da,接近中性的理论等电点为6.59。Prabi预测结果显示,MePOD10蛋白由α-螺旋、伸展链和不规则卷曲组成蛋白二级结构。MePOD10蛋白不稳定性指数为42.22,GRAVY指数(总平均疏水性)为−0.067,表明其是一种不稳定疏水性蛋白。WoLF PSORT II预测结果显示,MePOD10基因主要定位于细胞核膜上。NCBI-CDD数据库对MePOD10蛋白保守结构域预测,结果显示在24~324 bp区间存在一个POD保守结构域。2.2.2 多序列比对与进化分析
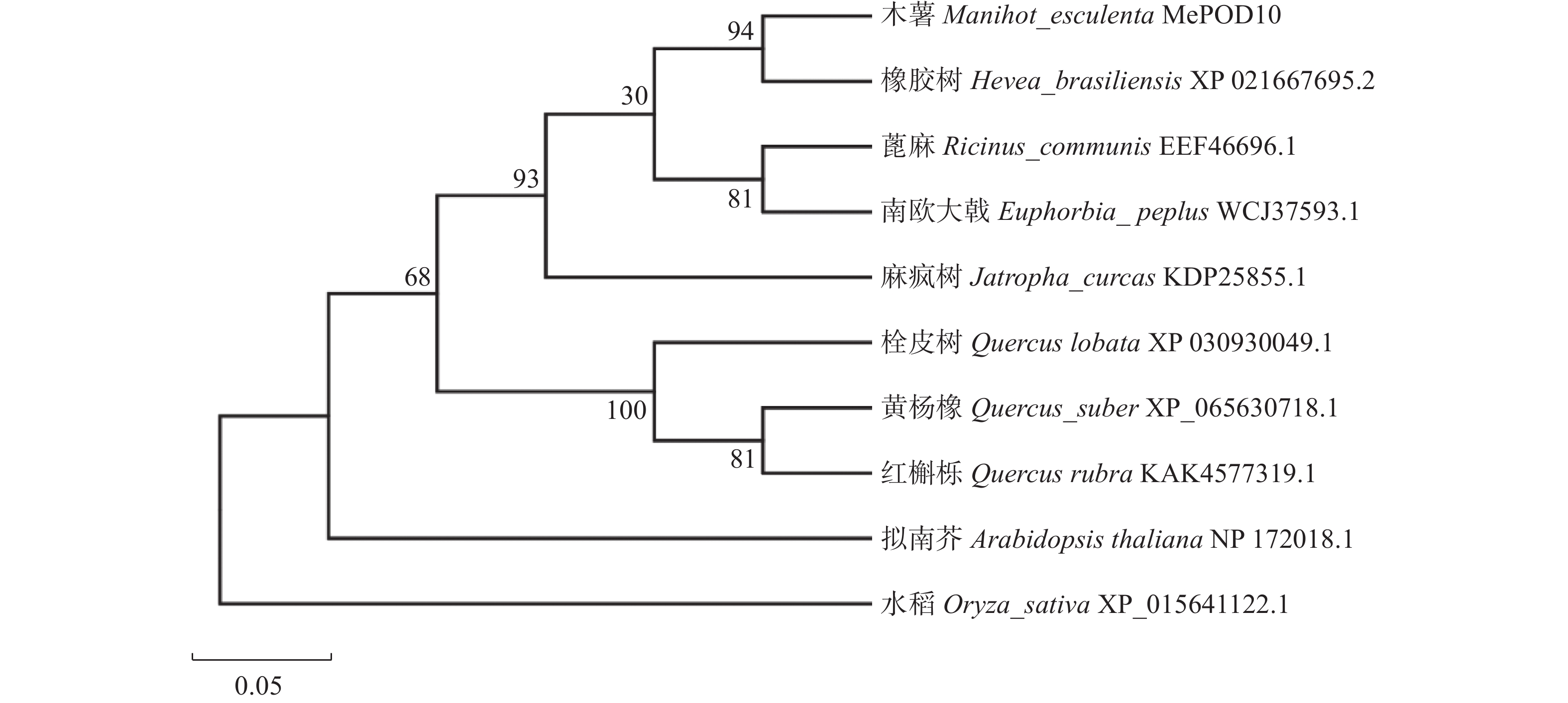
通过NCBI数据库中的BLAST工具对MePOD10的氨基酸序列进行比对分析,筛选与木薯MePOD10蛋白具有较高同源性的氨基酸序列,结果(图2)显示MePOD10与橡胶树(Hevea brasiliensis; XP_021667695.2)的POD蛋白氨基酸序列同源性最高,为93.25%。此外,与麻疯树(Jatropha curcas; KDP25855.1)和蓖麻(Ricinus communis; EEF46696.1)的同源性也较高,分别为88.04%和87.93%。系统发育进化树显示,MePOD10与橡胶树的POD蛋白亲缘关系最近(图3),表明两者在进化过程中保持了较高的序列相似性和功能保守性。
![]() 图 2 MePOD10与其他物种氨基酸序列同源性比对结果Figure 2. Alignment and homology of MePOD10 amino acid sequence
图 2 MePOD10与其他物种氨基酸序列同源性比对结果Figure 2. Alignment and homology of MePOD10 amino acid sequence2.3 MePOD10原核表达载体的构建及鉴定
利用同源重组方法将MePOD10基因构建于EcoRⅠ和BamHⅠ双酶切好的pET28a载体(图4A)上,再转化至DH5α感受态细胞中,涂板后挑取6个单克隆进行PCR验证,得到条带大小与目的片段一致的阳性克隆(图4B),将阳性单克隆摇菌后碱裂解法提取质粒,再双酶切质粒验证(图4C)。
![]() 图 4 MePOD10-pET28a载体构建M:DL2000 DNA标记,M1: 15000 DNA标记;A:1为pET28a空载,2为酶切pET28a;B:同源重组后菌落PCR;1~6为PCR检验;C:1~6为酶切6个MePOD10-pET28载体。Figure 4. Construction of MePOD10-pET28a vectorM: DL2000 DNA marker; M1: 15,000 DNA marker; A: 1: empty vector of pET28a; 2: enzyme-digested pET28a; B: PCR of homologous recombinant colonies; 1-6: PCR tests; C: 6 digested MePOD10-pET28a vectors.
图 4 MePOD10-pET28a载体构建M:DL2000 DNA标记,M1: 15000 DNA标记;A:1为pET28a空载,2为酶切pET28a;B:同源重组后菌落PCR;1~6为PCR检验;C:1~6为酶切6个MePOD10-pET28载体。Figure 4. Construction of MePOD10-pET28a vectorM: DL2000 DNA marker; M1: 15,000 DNA marker; A: 1: empty vector of pET28a; 2: enzyme-digested pET28a; B: PCR of homologous recombinant colonies; 1-6: PCR tests; C: 6 digested MePOD10-pET28a vectors.2.4 MePOD10重组蛋白的原核表达
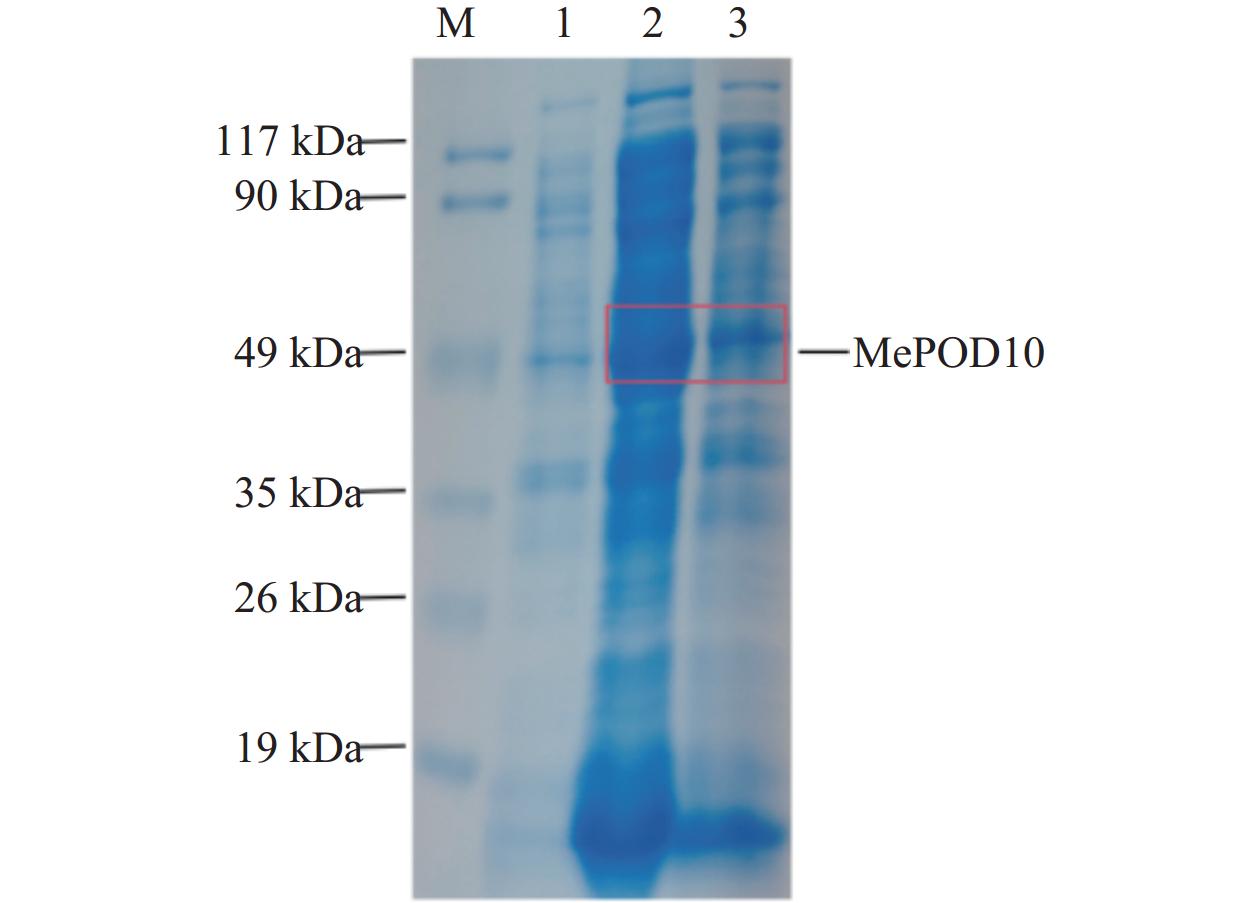
将验证正确的重组质粒转入BL21(DE3)感受态细胞中,在诱导温度为37 ℃,D600值为0.6~0.8条件下,加入IPTG至终浓度为1 mmol·L−1,并在200 r·min−1条件下培养。诱导蛋白表达6 h后,进行SDS-PAGE凝胶电泳检测。结果发现,在49 kDa之间有特异性条带,与预期的MePOD10重组蛋白大小一致(图5)。
![]() 图 5 MePOD10蛋白表达结果M:蛋白分子量标准;1:pET28蛋白;2:MePOD10蛋白上清;3:MePOD10蛋白沉淀。Figure 5. Expression of MePOD10M: Protein marker; 1: pET28a protein; 2: MePOD10 supernatant; 3: MePOD10 precipitation.
图 5 MePOD10蛋白表达结果M:蛋白分子量标准;1:pET28蛋白;2:MePOD10蛋白上清;3:MePOD10蛋白沉淀。Figure 5. Expression of MePOD10M: Protein marker; 1: pET28a protein; 2: MePOD10 supernatant; 3: MePOD10 precipitation.2.5 MePOD10重组蛋白的Western blotting验证
为进一步明确MePOD10重组蛋白表达的准确性,本研究利用Myc抗体,对收集的重组蛋白上清和沉淀进行Western blotting检测。结果表明:重组蛋白在49 kDa处出现特异性条带,说明重组蛋白表达成功(图6)。
![]() 图 6 蛋白Western blotting验证M:蛋白分子量标准;1:pET28蛋白;2:MePOD10蛋白上清;3:MePOD10蛋白沉淀。Figure 6. Protein validation by western blottingM: Protein marker; 1: pET28a protein; 2: MePOD10 supernatant; 3: MePOD10 precipitation.
图 6 蛋白Western blotting验证M:蛋白分子量标准;1:pET28蛋白;2:MePOD10蛋白上清;3:MePOD10蛋白沉淀。Figure 6. Protein validation by western blottingM: Protein marker; 1: pET28a protein; 2: MePOD10 supernatant; 3: MePOD10 precipitation.2.6 MePOD10重组蛋白的纯化及酶活性测定
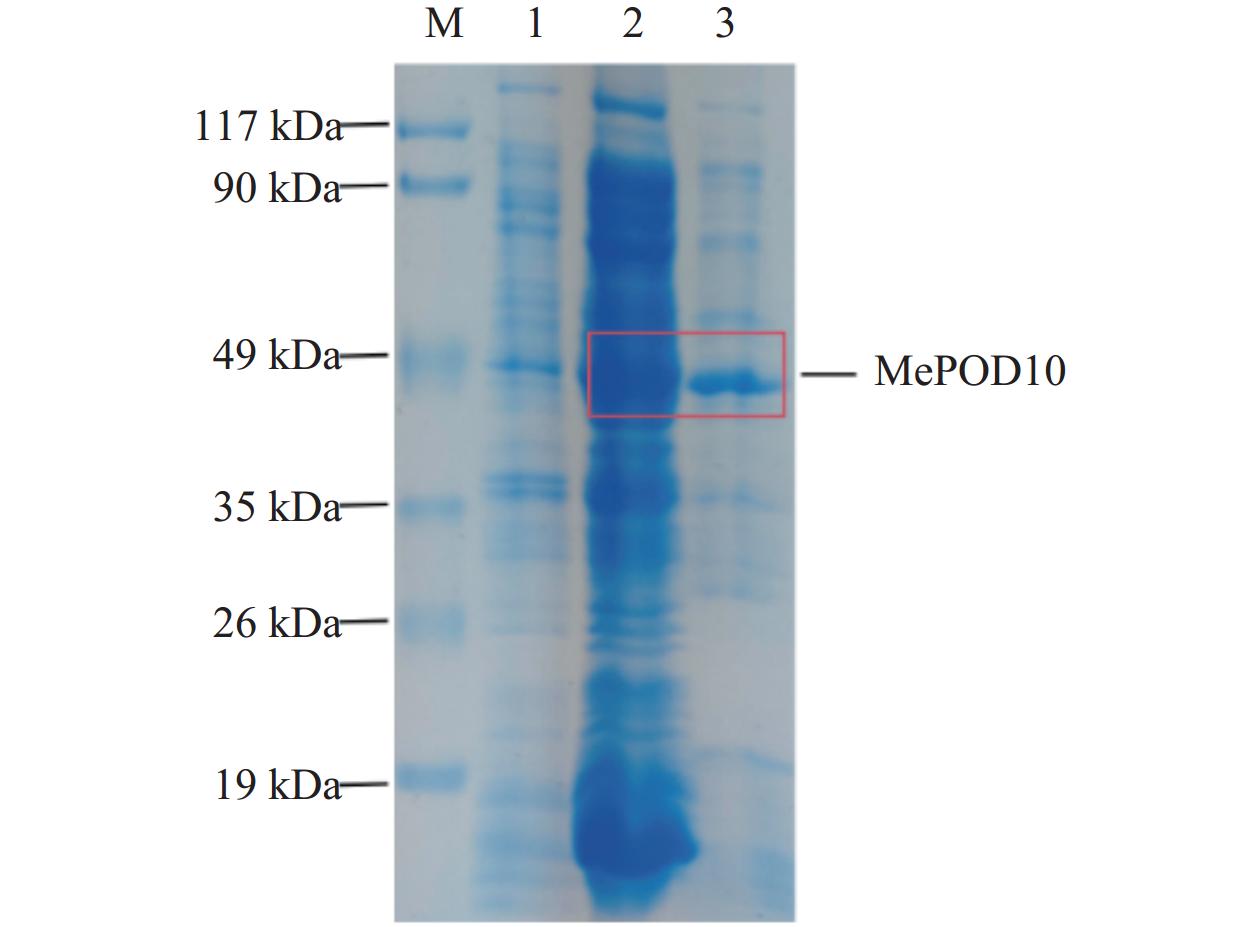
根据上述诱导条件对MePOD10重组蛋白大量诱导及纯化,获得纯化的蛋白(图7),大小与MePOD10重组蛋白大小一致,约49 kDa。由MePOD10重组蛋白的酶活性检测结果(图8A)可知,MePOD10蛋白酶活性显著高于对照蛋白(P<0.0001),说明MePOD10重组蛋白具有催化活性。以愈创木酚为底物的MePOD10蛋白酶动力学特性结果(图8B)表明,随着愈创木酚浓度增加,蛋白催化活性迅速增加,后趋于平稳,说明MePOD10蛋白具有POD酶活性。MePOD10蛋白Km值为3.113 mmol·L−1,表明MePOD10蛋白对愈创木酚亲和力大,酶促反应容易进行。
![]() 图 7 MePOD10上清蛋白纯化M:蛋白分子量标准;1:pET28蛋白;2:MePOD10蛋白上清;3:MePOD10纯化上清蛋白。Figure 7. Purification of MePOD10 supernatantM: protein marker; 1: pET28a protein; 2: MePOD10 supernatant; 3: purified MePOD10 supernatant.
图 7 MePOD10上清蛋白纯化M:蛋白分子量标准;1:pET28蛋白;2:MePOD10蛋白上清;3:MePOD10纯化上清蛋白。Figure 7. Purification of MePOD10 supernatantM: protein marker; 1: pET28a protein; 2: MePOD10 supernatant; 3: purified MePOD10 supernatant.![]() 图 8 MePOD10蛋白酶活性和动力学分析A:对照蛋白(pET28a)和MePOD10蛋白酶活测定;B:以愈创木酚为底物时,MePOD10蛋白酶学动力曲线。****表示差异显著(P<0.0001)。Figure 8. Enzymatic activity and kinetic analysis of MePOD10A: enzymatic activities of pET28a (control) and MePOD10; B: kinetics of MePOD10 protein observed with guaiacol as substrate. ****: significant difference at P<0.0001.
图 8 MePOD10蛋白酶活性和动力学分析A:对照蛋白(pET28a)和MePOD10蛋白酶活测定;B:以愈创木酚为底物时,MePOD10蛋白酶学动力曲线。****表示差异显著(P<0.0001)。Figure 8. Enzymatic activity and kinetic analysis of MePOD10A: enzymatic activities of pET28a (control) and MePOD10; B: kinetics of MePOD10 protein observed with guaiacol as substrate. ****: significant difference at P<0.0001.3. 讨论
随着全球气候的变化,木薯在生长发育的过程中受到各种胁迫与病害的制约,其中木薯花叶病是木薯最严重的病害之一。从经济、环保、稳定性和持久性等长远目光来看,常规分子育种是应对花叶病的有效长期策略[18]。
目前的木薯分子育种研究不仅挖掘到了针对CMD的单基因显性抗性CMD2基因座,还发现了另外两种抗性来源:多基因隐性抗性基因座CMD1以及与CMD2不关联的CMD3基因座。携带CMD2抗性位点的木薯几乎都表现出对木薯花叶病的高度抗性[19],随后有相关研究在CMD2基因座中找到了两个候选基因MePOD10和MePOD58,发现其与木薯花叶病的抗性紧密相关,但其具体的抗病功能还未得到验证。为进一步研究MePOD10在木薯中抗花叶病的功能,本研究构建了原核表达载体MePOD10-pET28a。原核表达载体有多种系列,其中pET是目前使用最为广泛的载体之一[20]。BL21(DE3)也是一种高效且成本较低的表达系统[21]。本研究所选用的pET28a载体和大肠杆菌 BL21(DE3)可以正确、大量地诱导MePOD10蛋白表达,该融合蛋白大小约为49 kDa,符合预测大小。此外,本研究发现在1 mmol·L−1 IPTG、37 ℃、200 r·min−1条件下诱导6 h可以得到较多的目的蛋白,且上清和沉淀均能表达出蛋白。相关研究表明影响大肠杆菌表达的因素有时间、温度等多方面,且37 ℃为大肠杆菌诱导的适宜温度[22]。
POD是植物体内一种非常重要且普遍存在的酶类。POD在植物体内扮演着多重角色,特别是在氧化还原反应和细胞保护方面发挥着关键作用,是植物体内的主要保护酶之一[23]。POD活性随植物生长发育进程以及环境条件的改变而变化,在逆境条件下(如干旱、盐胁迫、病虫害等),植物体内会产生大量的活性氧,导致氧化胁迫加剧。此时,POD等抗氧化酶的活性会显著增强,以清除植物体内过剩的活性氧,减轻氧化损伤的程度。因此,测定POD活性的高低可以作为衡量植物抗逆性强弱的一个重要指标。POD酶活性测定方法有多种,使用广泛且操作简便的方法是愈创木酚法[24]。在酶动力学研究中,Km值是酶特征性常数,可以代表酶与底物的亲和能力,且其大小与酶的性质有关,Km越小,表明酶与底物的亲和力越高,催化作用越大[25]。本研究以不同浓度的愈创木酚为底物时测定MePOD10活性,拟合酶动力学曲线发现,随着底物浓度增加,酶促反应逐渐上升,后趋于平稳,且Km值为3.113 mmol·L−1,表明MePOD10具有POD活性且对愈创木酚的亲和力和催化作用大。本研究初步测定MePOD10蛋白酶活性并拟合酶动力学曲线计算Km值,为进一步探究其蛋白功能奠定基础。
4. 结论
本研究对MePOD10基因进行克隆、原核表达分析以及蛋白酶活性和动力学分析研究,并且对其进行了生物信息学和原核表达情况的初步分析,探索了一些目的蛋白的诱导方法和条件,具有一定的前瞻性和创新性。由于目的蛋白在上清表达量相对沉淀较少,在进一步纯化蛋白试验中目的蛋白纯度较低,因此诱导目的蛋白的更优条件仍有待深入发掘。
-
![]()
图 1 PpCaMK1核心片段的克隆(A)和PpCaMK1 3′端和5′端的克隆(B)
M:DL1000 DNA Marker。 A:1~3分别为核心片段1、核心片段2、核心片段3 。B:1为3′RACE ,2为5′RACE。
Figure 1. Cloning PpCaMK1 core fragment (A) and 3' and 5' (B)
M: DL1000 DNA marker; A: 1, 2, and 3 were core fragments 1, 2, and 3, respectively; B: 1 was 3′RACE, 2 was 5′RACE.
![]()
图 2 水滴伪康纤虫CaMK1基因全长cDNA序列及对应的氨基酸序列
ORF用大写字母表示,5′-UTR和3′-UTR用小写字母表示。起始密码子加框显示,终止密码子以星号标注。
Figure 2. Full-length cDNA sequence and corresponding amino acid sequence of PpCaMK1
ORF domain shown in uppercase letters; 5′-UTR and 3′-UTR domains in lowercase letters; start codon in box; stop codon with asterisk.
![]()
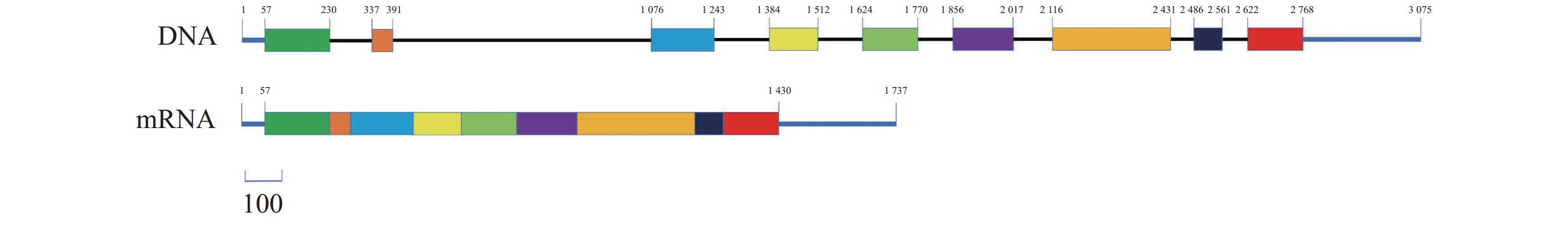
图 3 水滴伪康纤虫CaMK1的DNA和mRNA结构
蓝线表示非翻译区,黑线表示内含子,长方形表示外显子。
Figure 3. Schematic DNA and mRNA of PpCaMK1
Blue line represents untranslated region; black line, introns; and rectangles, exons.
![]()
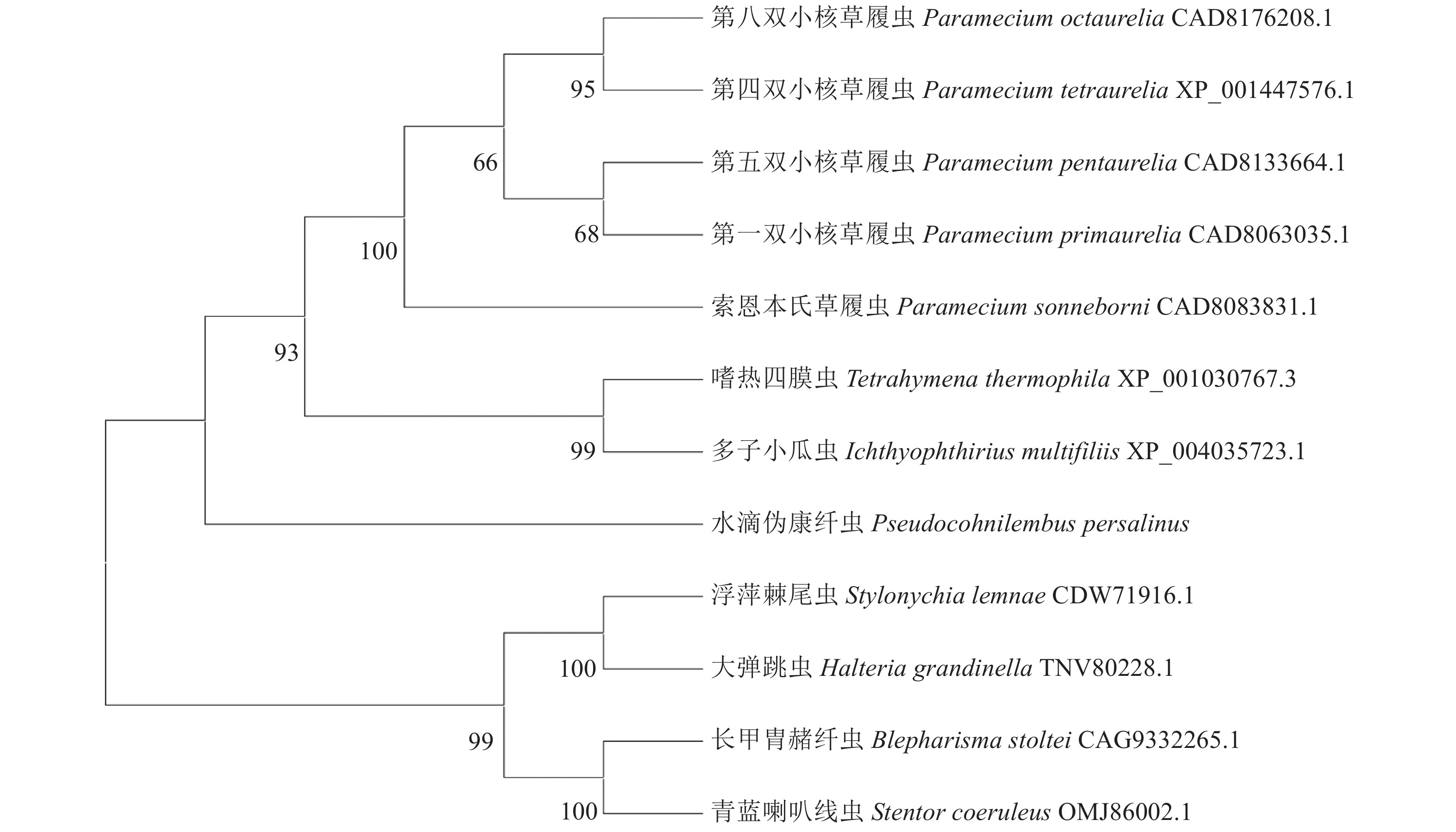
图 6 水滴伪康纤虫PpCaMK1蛋白与其他生物的多序列比对
Figure 6. Multiple sequences of PpCaMK1 and those of other organisms
表 1 水滴伪康纤虫CaMK1基因克隆中应用的引物
Table 1 Primers used for cloning PpCaMK1
引物名称
Primer name引物序列(5′-3′)
Primer sequence(5′-3′)目的
Purposec15077 F1 GGTTTAGAGTTTAGACGCTGA PpCaMK1核心片段1克隆 c15077 R1 CTTCGTCCCAGTCGATTAAC PpCaMK1核心片段1克隆 c15077 F2 TTGGTGTTTCTATGGTCGCA PpCaMK1核心片段2克隆 c15077 R2 CTTCTTCGTTTACCTCCTCC PpCaMK1核心片段2、3克隆 c15077 F3 GCTGATGATGGACAGTTAATCG PpCaMK1核心片段3克隆 c15077 3′ F1 GATTACGCCAAGCTTCGACTGGGACGAAGAGGATTCAAAGTGG 第一轮3′ RACE 克隆 c15077 3′ F2 GATTACGCCAAGCTTGTTCAAAGGGTTACTCTGGTAAAGCTGC 第二轮3′ RACE克隆 c15077 5′ R1 GATTACGCCAAGCTTCCACTTTGAATCCTCTTCGTCCCAGTCG 第一轮5′ RACE克隆 c15077 5′ R2 GATTACGCCAAGCTTTTACCATCAGCGTCTAAACTCTAAACCAT 第二轮5′ RACE克隆 UPM(long) SMARTer RACE 5′/3′Kit(TAKARA code No.050421)提供 通用长引物 UPM(short) SMARTer RACE 5′/3′Kit(TAKARA code No.050421)提供 通用短引物  下载: 导出CSV
下载: 导出CSV
-
[1] 常志强,陈钊,李泽鹏,等. 中兽药驱虫复方制剂防治海水养殖中盾纤毛虫的临床试验研究[J]. 中兽医医药杂志,2019,38(5) :5−9. CHANG Z Q,CHEN Z,LI Z P,et al. Clinical assessment of traditional Chinese medicine compound on the ciliate parasite in marine aquaculture[J]. Journal of Traditional Chinese Veterinary Medicine,2019,38(5) :5−9. (in Chinese)
[2] IGLESIAS R,PARAMÁ A,ALVAREZ M F,et al. Philasterides dicentrarchi (Ciliophora,Scuticociliatida) as the causative agent of scuticociliatosis in farmed turbot Scophthalmus maximus in Galicia (NW Spain) [J]. Diseases of Aquatic Organisms,2001,46(1) :47−55.
[3] 崔龙波,刘冉,王琛,等. 大菱鲆盾纤毛虫病的流行病学调查[J]. 科学养鱼,2015(3) :54−56. CUI L B,LIU R,WANG C,et al. Epidemiological investigation of Scophthalmus maximus shield ciliary disease[J]. Scientific Fish Farming,2015(3) :54−56. (in Chinese)
[4] 潘旭明,马洪钢,邵晨,等. 海洋纤毛虫:水滴伪康纤虫的口器发生及形态学重描述[J]. 水生生物学报,2012,36(3) :489−495. DOI: 10.3724/SP.J.1035.2012.00489 PAN X M,MA H G,SHAO C,et al. Stomatogenesis and morphological redescription of Pseudocohnilembus persalinus (Ciliophora:Scuticociliatida) [J]. Acta Hydrobiologica Sinica,2012,36(3) :489−495. (in Chinese) DOI: 10.3724/SP.J.1035.2012.00489
[5] BRAUN A P,SCHULMAN H. The multifunctional calcium/calmodulin-dependent protein kinase:From form to function[J]. Annual Review of Physiology,1995,57:417−445. DOI: 10.1146/annurev.ph.57.030195.002221
[6] 肖圣燕,程隆基,许凯,等. 桑褐斑壳丰孢菌pmcamk基因的克隆与表达时相分析[J]. 蚕业科学,2023,49(1) :15−22. XIAO S Y,CHENG L G,XU K,et al. Cloning and expression analysis of calcium /calmodulin-dependent protein kinases gene of Phloeospora maculans[J]. Acta Sericologica Sinica,2023,49(1) :15−22. (in Chinese)
[7] 吴迪,赵梓彤,李志勇,等. 玉米大斑病菌钙/钙调素依赖性蛋白激酶(CaMK) 基因家族的鉴定与表达模式分析[J]. 农业生物技术学报,2015,23(8) :1020−1030. WU D,ZHAO Z T,LI Z Y,et al. Identification and expression pattern analysis of calcium/calmodulin dependent protein kinases (CaMKs) gene family in Setosphaeria turcica[J]. Journal of Agricultural Biotechnology,2015,23(8) :1020−1030. (in Chinese)
[8] 蒋倩倩,毛仁燕,李永才,等. 梨果黑斑病菌AaCaMK基因克隆、生物信息学分析及其在侵染结构分化中的表达分析[J]. 微生物学通报,2021,48(12) :4664−4676. JIANG Q Q,MAO R Y,LI Y C,et al. Cloning,bioinformatics and expression analysis of AaCaMK gene on infection structure differentiation of Alternaria alternate,causal agent of pear black spot[J]. Microbiology China,2021,48(12) :4664−4676. (in Chinese)
[9] KATO K,SUDO A,KOBAYASHI K,et al. Characterization of Plasmodium falciparum protein kinase 2[J]. Molecular and Biochemical Parasitology,2008,162(1) :87−95. DOI: 10.1016/j.molbiopara.2008.07.007
[10] KATO K,SUGI T,TAKEMAE H,et al. Characterization of a Toxoplasma gondii calcium calmodulin-dependent protein kinase homolog[J]. Parasites &Vectors,2016,9(1) :405.
[11] XIONG J,WANG G Y,CHENG J,et al. Genome of the facultative scuticociliatosis pathogen Pseudocohnilembus persalinus provides insight into its virulence through horizontal gene transfer[J]. Scientific Reports,2015,5:15470.
[12] 林能锋,曾红. 水滴伪康纤虫全长cDNA文库的构建及EST分析[J]. 福建农业学报,2017,32(9) :921−925. LIN N F,ZENG H. Full-length cDNA Library and EST Analysis on Pesudocohnilembus persalinus[J]. Fujian Journal of Agricultural Sciences,2017,32(9) :921−925. (in Chinese)
[13] 贺斌,黄兴奇,余腾琼,等. cDNA末端快速扩增技术及其方法的改进[J]. 浙江农业科学,2012,53(9) :1352−1357. DOI: 10.3969/j.issn.0528-9017.2012.09.045 HE B,HUANG X Q,YU T Q,et al. Rapid amplification technology of cDNA terminal and its improvement[J]. Journal of Zhejiang Agricultural Sciences,2012,53(9) :1352−1357. (in Chinese) DOI: 10.3969/j.issn.0528-9017.2012.09.045
[14] HALL T A. BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT[J]. Nucleic Acids Symposium Series,1999,41(2) :95−98.
[15] KUMAR S,TAMURA K,NEI M. MEGA:Molecular Evolutionary Genetics Analysis software for microcomputers[J]. Computer Applications in the Biosciences,1994,10(2) :189−191.
[16] 宫春光,李凤晨. 牙鲆盾纤毛虫病的药物防治试验[J]. 水产科学,2007,26(9) :509−511. DOI: 10.3969/j.issn.1003-1111.2007.09.009 GONG C G,LI F C. Chemical treatment on protozoon Paralembus digitiformis in farmed Japanese flounder[J]. Fisheries Science,2007,26(9) :509−511. (in Chinese) DOI: 10.3969/j.issn.1003-1111.2007.09.009
[17] 林能锋,郑晨艳,王侯杰,等. 中草药浸出液体外驱杀两种水产致病性盾纤虫的研究[J]. 福建畜牧兽医,2017,39(5) :13−15. DOI: 10.3969/j.issn.1003-4331.2017.05.006 LIN N F,ZHENG C Y,WANG H J,et al. Repellent effect of Chinese herbal leachate to two species of scuticociliate in vitro[J]. Fujian Journal of Animal Husbandry and Veterinary Medicine,2017,39(5) :13−15. (in Chinese) DOI: 10.3969/j.issn.1003-4331.2017.05.006
[18] 崔青曼,袁春营,李春岭,等. 主要海水养殖鱼类白点病和盾纤毛虫病防治技术[J]. 水利渔业,2007,27(6) :85−87,94. DOI: 10.3969/j.issn.1003-1278.2007.06.037 CUI Q M,YUAN C Y,LI C L,et al. Prevention and control technology of white spot disease and scuticociliatosis in major marine aquaculture fish[J]. Journal of Hydroecology,2007,27(6) :85−87,94. (in Chinese) DOI: 10.3969/j.issn.1003-1278.2007.06.037
[19] 刘荣荣,张进顺,张晓磊,等. 刚地弓形虫棒状体蛋白10的生物信息学分析[J]. 中国病原生物学杂志,2017,12(9) :855−859. LIU R R,ZHANG J S,ZHANG X L,et al. Bioinformatic analysis of rhoptry protein 10 of Toxoplasma gondii[J]. Journal of Pathogen Biology,2017,12(9) :855−859. (in Chinese)
[20] SIVALINGAM G N,SHEPHERD A J. An analysis of B-cell epitope discontinuity[J]. Molecular Immunology,2012,51(3/4) :304−309.
[21] HARMON A C,GRIBSKOV M,HARPER J F. CDPKs–a kinase for every Ca2+ signal?[J]. Trends in Plant Science,2000,5(4) :154−159. DOI: 10.1016/S1360-1385(00)01577-6
[22] KLIMECKA M,MUSZYŃSKA G. Structure and functions of plant calcium-dependent protein kinases[J]. Acta Biochimica Polonica,2007,54(2) :219−233. DOI: 10.18388/abp.2007_3242
[23] LIU H L,CHE Z J,ZENG X R,et al. Genome-wide analysis of calcium-dependent protein kinases and their expression patterns in response to herbivore and wounding stresses in soybean[J]. Functional &Integrative Genomics,2016,16(5) :481−493.
[24] 费小钰,李红丽,王俊皓. 植物钙依赖蛋白激酶CDPK基因功能综述[J]. 吉林农业,2017(9) :104−105. FEI X Y,LI H L,WANG J H. Review on the function of plant calcium-dependent protein kinase CDPK gene[J]. Agriculture of Jilin,2017(9) :104−105. (in Chinese)
[25] SWULIUS M T,WAXHAM M N. Ca2+/calmodulin-dependent protein kinases[J]. Cellular and Molecular Life Sciences,2008,65(17) :2637−2657. DOI: 10.1007/s00018-008-8086-2








计量
- 文章访问数: 30
- HTML全文浏览量: 3
- PDF下载量: 4
