

IL15RA Mutation Site and Relationship with Growth of Goats
-
摘要:目的
检测福清山羊和努比亚山羊群体细胞介素15受体亚基α(IL15RA–interleukin 15 receptor subunit alpha, IL15RA)基因的表达谱以及突变位点多态性,分析不同位点与山羊生长性状的相关性,进一步分析IL15RA基因对山羊生长性状的影响。
方法随机挑选福清山羊和努比亚山羊12月龄羯羊各3只,分别采集心、肝、脾、肺、肾、头颈半棘肌、背最长肌、股四头肌和臀股二头肌共9种组织,采用qRT-PCR方法构建该基因在两个品种不同组织中的表达谱,Sanger测序检测福清山羊(n=108)和努比亚山羊(n=286)群体中IL15RA基因的SNP位点,并分析其与生长性状的相关性。
结果(1)IL15RA基因在肌肉组织中的表达水平显著低于内脏组织(P<0.05),福清山羊背最长肌表达水平显著低于努比亚山羊(P<0.05),肺部组织表达量显著高于努比亚山羊(P<0.05)。(2)福清山羊和努比亚山羊IL15RA基因内含子3存在1个InDel位点,为g.22224_
22225 Del A,Del22224。内含子6存在1个SNP位点,为g.33428A>G。(3)SNP-g.33428位点显著影响(P<0.05)福清山羊的体重、胸围、胸宽、胸深、尻宽等指标。(4)在福清山羊中,单倍型组合DD/AA、ID/GG为优势单倍型组合;在努比亚山羊中,单倍型组合ID/AA、II/GG为优势单倍型组合。结论IL15RA基因多态性与福清山羊和努比亚山羊生长性状显著相关,IL15RA基因可作为福清山羊和努比亚山羊生长性状选育的候选基因。
Abstract:ObjectiveThe interleukin 15 receptor subunit α gene (IL15RA) in goats was studied to identify the mutation site that differentiates the growth of Fuqing and Nubian goats.
MethodsThree 12-month-old Fuqing and Nubian goats each were randomly selected to collect tissue samples of the heart, liver, spleen, lung, and kidney as well as the head and neck semispinalis muscles, longest back muscle, thigh muscle, and biceps brachii muscle for qRT-PCR to construct a spectrum of expression variations between the two goat species. The SNP of IL15RAs extracted from the Fuqing goats (n=108) and the Nubian goats (n=286) were subjected to a PCR hybridization Sanger sequencing for genotype classification using PCR-RFLP and correlation with goat growth.
Results(1) The IL15RA expressions in the muscles were significantly lower than those in the viscera (P<0.05); the expression in the longest back muscle of the Fuqing goats significantly lower than that of the Nubian goats (P<0.05); and the expression in the lung of the Fuqing goats significantly higher than that of the Nubian goats (P<0.05). (2) One InDel site, g.22224_222225 Del A, Del22224, was found in the intron 3 of IL15RAs of Fuqing and Nubian goats, and one SNP site, g.33428A>G, in the intron 6. (3) SNP-g.33428 significantly affected the body weight, chest circumference, chest width, and hucklebone width of Fuqing goats (P<0.05). (4) DD/AA and ID/GG were the dominant haplotype combinations in the Fuqing goats. It was ID/AA and II/GG in the Nubian goats.
ConclusionThe polymorphisms of IL15RAs significantly related to the differentiate growth traits of Fuqing and Nubian goats. The gene could conceivably become the target for improving the currently practiced molecular markers-assisted breeding programs.
-
Keywords:
- goat /
- IL15RA /
- single nucleotide polymorphisms /
- growth and development /
- correlation analysis
-
0. 引言
【研究意义】玉米是我国重要的粮食作物,玉米种子表面携有大量微生物,这些微生物有可能是影响玉米种子出芽、出苗的病原菌,也有可能是田间作物生长期侵染性致病菌,如引起根茎腐病、鞘腐病、穗腐病、顶腐病、丝黑穗病发生的病原真菌和细菌[1]。为了提高玉米种子的出芽率,避免播种后种子携带的病原菌进入土壤,在玉米生长季节引发病害发生,采用合理的方式进行玉米种子表面微生物广谱性消毒处理具有重要的意义。【前人研究进展】当前,对种子常见的消毒方法主要有采用福尔马林、次氯酸钠、农用链霉素、苯醚甲环唑等化学消毒剂或杀菌剂处理[2-5],而采用化学杀菌剂浸种或拌种是玉米种子杀菌消毒处理最常用的技术,已有研究表明采用苯醚甲环唑浸种处理能预防玉米真菌性病害丝黑穗病、茎基腐病发生[5],但药剂浸种处理靶标性强,对药剂浓度、剂型、药种比、时间要求严格,使用不当易引起药害,不利于出苗或效果不佳。酸性氧化电位水(Acid electrolyzed oxidizing water,AEOW),简称酸化水,是电解水中酸性最强、杀菌率最高的一种新型广谱、高效、环保消毒剂[6]。该酸化水系普通自来水中加入低浓度氯化钠(溶液浓度小于0.1%),经电解处理后,从阳极区生成具有低浓度有效氯(50~102 ppm)、高氧化还原电位(ORP值≥1100 mV)和低pH值(pH 2.0~3.0)的酸性水溶液,其成分主要为氯气、次氯酸、盐酸等[7]。其杀菌机理主要是低pH值可破坏病原微生物的生存环境,H+离子透过微生物细胞膜,间接参与微生物的代谢过程,导致杀灭、抑制微生物;而高氧化还原电位值则影响微生物的呼吸和能量代谢,改变病原微细胞膜电位,增强膜的透性,导致细胞肿胀崩解,使微生物迅速死亡;酸化水中的有效氯也具有直接杀菌作用,上述3个杀菌因子的协同联合作用使酸化水能达到快速杀灭病原微生物的目的[8-9]。酸化水最先用于医药领域,经多年的研究、实践,人们对其杀菌的有效性、安全性、不留残毒、有利于环保等优点形成共识,现已在医疗领域广泛应用[10-13]。酸化水也常作为绿色杀菌剂应用于食品、环保、农业等行业[8,14],对果蔬、肉类和水产品具有防腐保鲜、降解农残等功效,在食品加工上也具有极大的应用前景[15-17]。在农业上,目前主要用于防治作物病害和改良土壤[18-20],有研究表明,喷施酸化水对小麦赤霉病[20],黄瓜白粉病[21-22],黄瓜霜霉病[18,23],稻曲病[24]具有较好的防病效果。【本研究切入点】因酸化水在田间病害防治应用上存在防治次数多、用液量大、使用成本高等问题,限制了酸化水在农业植物病害防治中的推广。然而,关于利用酸化水对作物种子表面微生物消毒作用有待进一步探讨。【拟解决的关键问题】我国是一个农业大国,农业生产过程中农药使用量大,为拓展酸化水在农业生产中的应用领域,探索其对玉米种子表面微生物或病原菌的杀菌效果,预防种传病害发生的风险性,本研究测定并评价了酸化水对玉米种子表面微生物的抑制作用及其对玉米种子出芽和出苗的安全性,旨在为农业生产实践中合理使用酸化水和减少农药用量提供参考。
1. 材料与方法
1.1 酸性氧化电位水
酸性氧化电位水由杭州美美科技有限公司生产提供(有效氯含量:90 mg·L−1,氧化还原电位>1 100 mv,pH值:3)。
1.2 玉米品种
福建省主栽鲜食玉米品种泰鲜甜1号和泰鲜甜2号,由福建省圆旺园种子集团有限公司提供。
1.3 病原菌菌株
玉米鞘腐病病原菌拟轮枝镰孢菌PNQF18(Fusarium verticillium)和玉米茎腐病病原菌假单孢杆菌MHXJ01(Pseudomonas spp.)由福建省农业科学院植物保护研究所提供。
1.4 试验方法
1.4.1 酸化水不同处理对玉米种子表面离体微生物的抑制作用
取泰鲜甜1号、泰鲜甜2号种子各15粒[百粒重(12.5 ±0.6) g]放入装有30 mL无菌水的灭菌离心管(50 mL)中, 28 ℃、80 r·min−1振荡6 h,制备种子表面微生物菌悬液。吸取2个品种菌悬液各1 mL,与酸化水依次按体积比1︰1、1︰2、1︰5、1︰10的比例均匀混合于灭菌的试管(15 mm×150 mm)内,分别静置0.5、1.0、2.0、4.0、6.0 h后,每处理各吸取0.20 mL混合液均匀涂布于直径9 cm的PDA平板上,无菌条件下吹干培养基表面多余水分,置于20 ℃条件下培养2 d,计数平板菌落数,计算每mL菌悬液中的含菌量,以无菌水代替酸化水,即将菌悬液与无菌水按体积比1︰10的比例混合后的液体静置6 h为空白对照。每处理共检测3皿,试验重复3次。
1.4.2 酸化水不同处理对玉米种子带菌率的影响
取供试种子各50粒于灭菌的三角瓶中,分别加入酸化水50、75和100 mL,使种子与酸化水依次按质量体积比(mg/mL)125︰1、125︰1.5和125︰2,28 ℃、80 r·min−1分别振荡处理1、2、4、6 h后,将种子移入PDA平板上,25 ℃黑暗条件下培养5 d,检测平板种子带菌情况。以用100 mL无菌水代替酸化水振荡处理种子6 h为空白对照,试验重复3次。另从酸化水3种用量处理1、2、4、6 h的种子中,等量抽样调查150粒种子携带细菌或真菌情况,统计酸化水不同处理时间各类菌株出现频次和出现频率(同时出现细菌和真菌的个别种子归为携带细菌的种子)。种子带菌率/%= (带菌种子数/供试种子数)×100;菌株出现频率/%= (菌株出现的种子数/带菌种子数)×100。
1.4.3 酸化水不同处理对玉米种子出芽和胚根生长的影响
试验分别设为病原真菌预先接种组、病原细菌预先接种组和未接种组。参照田艳丽[25] 的方法进行玉米种子外源病菌接种处理。取泰鲜甜1号、泰鲜甜2号种子各300粒于灭菌的三角瓶中,分别用150 mL拟轮枝镰孢菌菌液(1×108个孢子·mL−1)和假单胞杆菌菌液(1×1010 cfu·mL−1)于20 ℃,80 r·min−1振荡浸泡处理2 h,制备病原真菌和细菌预先接种的玉米病种,以无菌水替代病菌菌液为玉米未接种组。另从试验的3组中,每组取泰鲜甜1号、泰鲜甜2种子各50粒,分别放入装有100 mL酸化水的灭菌三角瓶中,参照1.4.2的方法采用酸化水处理4 h和6 h后,取出各处理种子放入75%酒精擦拭过并铺设湿润无菌滤纸的方形塑料盒内(30 cm×20 cm×15 cm),于22 ℃黑暗条件下培育5 d后,观察种子出芽情况,计算出芽率(以芽长高于0.4 cm视为出芽)。从各处理出芽的种子中随机测量10~15粒种子芽长、主胚根长,计数胚根数。以用等量的无菌水替代酸化水处理种子6 h为空白对照,试验重复3次。出芽率/%= (出芽种子数/供试种子数)×100。
1.4.4 酸化水处理对不同玉米品种出苗和植株生长的影响
取泰鲜甜1号、泰鲜甜2号种子各50粒,分别放入装有100 mL酸化水的三角瓶中,参照1.4.2的方法采用酸化水处理6 h后,取出种子播种于含营养基质的育苗盘中,每穴播种1粒种子,于(22±2 )℃温室条件下育苗16 d,观察种子出苗情况,计算出苗率(以苗高于2 cm视为出苗)。从各处理出苗的植株中随机选取10~15株玉米苗,测量其株高(茎基部至次新叶叶鞘基部),并对其地上部鲜重进行称重。以用等量的无菌水替代酸化水处理种子6 h为空白对照,试验重复3次。出苗率/%=(出苗种子数/供试种子数)×100。
1.5 数据统计及分析
试验数据采用Excel 2007和DPS 7.5软件进行统计与分析。
2. 结果与分析
2.1 酸化水不同处理对玉米种子表面微生物的抑制作用
泰鲜甜1号和泰鲜甜2号种子表面含有大量的微生物,其15粒种子与30 mL无菌水混匀后,获得的微生物菌悬液含菌量均高于10 000 cfu·mL−1。酸化水4种不同用量和5种时间处理组合的抑菌效果如表1所示,泰鲜甜1号、泰鲜甜2号种子菌悬液与酸化水按(mL/mL)1︰5和1︰10比例混匀0.5~6.0 h后,检出的菌量均低于2 cfu·mL−1,而与酸化水按1︰1比例混匀0.5~6.0 h后,检出的菌量分别在7.67~100.44 cfu·mL−1和2.00~16.11 cfu·mL−1。酸化水处理0.5 h和1.0 h,2个品种菌悬液与酸化水按1︰5和1︰10比例混匀后,检出的菌量显著少于两者按1︰1和1︰2比例处理的菌量,而在1︰1比例处理的2 、4、6 h混合液中检出的菌量显著少于0.5 h和1.0 h混合液中的菌量。表明酸化水处理对玉米种子表面微生物有抑制作用,菌悬液与酸化水按不同比例混匀后,随着酸化水用量增加及作用时间的延长,检测出的菌量越少,采用酸化水2倍用量处理种子表面微生物菌悬液6 h便可达到较好的抑菌效果。
表 1 酸化水不同处理对玉米种子表面微生物菌悬液含菌量的影响Table 1. Effects of AEOW treatment on microbial count of maize seed surface rinse品种
Varieties时间
Time/h菌量 colony amount (cfu·mL−1) 菌悬液︰酸化水
MS︰AEOW 1︰1菌悬液︰酸化水
MS︰AEOW 1︰2菌悬液︰酸化水
MS︰AEOW 1︰5菌悬液︰酸化水
MS︰AEOW 1︰10对照
Control泰鲜甜1号 Taixiantian NO.1 0.5 100.44±9.96 Aa 17.78±1.07 Dd 0.67±0.67 Gf 0.44±0.51 Gf — 1 73.22± 3.08 Bb 14.89±3.24 DEd 0.22±0.19 Gf 0.00± 0.00 Gf — 2 29.11± 6.71 Cc 2.00± 0.88 Gf 0.00 ±0.00 Gf 0.00 ±0.00 Gf — 4 9.56± 2.14 EFe 0.67± 0.33 Gf 0.00 ±0.00 Gf 0.00 ±0.00 Gf — 6 7.67± 2.00 FGe 0.44± 0.73 Gf 0.00± 0.00 Gf 0.00 ±0.00 Gf >10000 泰鲜甜2号 Taixiantian NO.2 0.5 16.11± 4.67 Aa 10.00± 5.24 Bb 1.22±0.84 DEde 0.89± 0.51 Ede — 1 9.00±0.58 BCb 5.33± 1.20 CDc 0.44± 0.51 Ede 0.00± 0.00 Ee — 2 5.33± 0.67 CDc 2.33 ±1.53 DEcde 0.00± 0.00 Ee 0.00 ±0.00 Ee — 4 3.44± 1.58 DEcd 1.44± 0.51 DEde 0.00± 0.00 Ee 0.00 ±0.00 Ee — 6 2.00± 1.00 DEde 0.00± 0.00 Ee 0.00± 0.00 Ee 0.00±0.00 Ee >10000 ①不同大小写字母表示经Duncan氏新复极差法检验在 P<0.01和P<0.05水平差异显著;表2~6同。②MS:菌悬液;AEOW:酸化水.
① Data with different capital letters and lowercase letters on same column indicate significant differences at P<0.01 and P<0.05, respectively, according to Duncan's new multiple range method. The same as Table 2-6. ②MS:Microbial suspension; AEOW: Acid electrolyzed oxidizing water.2.2 酸化水不同处理对玉米种子带菌率的影响
未经酸化水处理的泰鲜甜1号和泰鲜甜2号种子带菌率均为100%。将种子与酸化水按不同用量比例混匀后,酸化水3种用量和4种处理时间组合的抑菌效果如表2所示,在酸化水相同时间处理下,2个品种种子与酸化水按(mg/mL)125︰2比例混匀后,种子带菌率显著低于两者按125︰1.5和125︰1比例的处理,而在酸化水相同用量下,试验的4种处理时间,均以酸化水处理6 h时的种子带菌率最低。其中泰鲜甜1号、泰鲜甜2号种子与酸化水按125︰2比例浸泡振荡处理6 h,检出的种子带菌率分别为14.00%和7.33%,显著低于酸化水另两种用量处理1 、2 、4 h的种子带菌率。表明酸化水处理对玉米种子有较好的表面消毒作用,酸化水用量增加及作用时间的延长,其消毒效果越好。
表 2 酸化水不同处理对玉米种子带菌率的影响Table 2. Effect of AEOW treatment on bacteria-retention rate of maize seeds处理 Treatment 种子带菌率 Seed infection rate/% 种子Seed∶酸化水
AEOW (mg)∶(mL)时间
Time/h泰鲜甜1号
Taixiantian NO.1泰鲜甜2号
Taixiantian NO.2125∶1 1 87.00±7.00 Ab 73.33±5.03 Bb 2 63.67±4.73 BCc 48.67±4.16 Dd 4 46.00±6.00 DEd 32.00±4.00 Ee 6 32.00±7.21 EFf 14.67±4.16 GHg 125∶1.5 1 69.33±7.57 Bc 62.67±6.11 Cc 2 44.67±4.16 DEd 34.67±7.02 Ee 4 43.33±3.06 DEde 22.00±3.46 FGf 6 24.67±7.02 FGf 10.00±2.00 Hg 125∶2 1 52.00±7.21 CDd 50.00±5.29 Dd 2 34.00±9.17 EFef 28.67±3.06 EFef 4 28.67±4.16 Ff 9.33±2.31 Hg 6 14.00±3.46 Gg 7.33±2.31 Hg 对照 Control 6 100.00±0.00 Aa 100.00±0.00 Aa 表 3 酸化水不同处理时间对玉米种子真菌和细菌出现频率的影响Table 3. Effects of AEOW treatment time on bacterial and fungal occurrence frequency on seeds时间
Time/h出现频率 Frequent/% 细菌
Bacteria真菌
Fungi镰孢菌
Fusarium曲霉/青霉
Aspergillus/Penicillium其他真菌
Other fungi1 15.33 50.00 34.67 12.00 3.33 2 9.33 32.67 23.33 8.00 1.33 4 4.67 25.33 20.00 4.00 1.33 6 1.33 20.00 16.00 3.33 0.67 表 4 酸化水不同处理对泰鲜甜1号种子出芽和胚根生长的影响Table 4. Effects of AEOW treatment on seed germination and radicle growth of Taixiantian No. 1组别
Group时间
Time/h出芽率
Germination rate/%芽长
Shoot length/cm主胚根长度
Radicle length/cm胚根数
No of primary root未接种 Without inoculation 4 71.33±5.03 Aa 3.65±0.64 Aa 9.71±0.63 Aa 5.27±0.24 Aa 6 73.33±7.02 Aa 3.69±0.23 Aa 9.85±0.42 Aa 4.92±0.21 ABCab 对照 Control 60.00 ±4.00 ABCbc 3.38±0.14 ABab 9.12±0.32 ABa 4.76±0.14 ABCbc 真菌接种 Fungus inoculation 4 66.67±4.16 ABab 2.84±0.16 Bbc 8.15±0.35 BCb 4.63±0.15 BCbc 6 68.37±5.03 ABab 3.22±0.06 ABabc 9.45±0.37 Aa 5.16±0.15 ABa 对照 Control 51.33±7.02 Cc 2.68±0.52 Bc 7.56±0.68 CDbc 4.56±0.12 Cbc 细菌接种 Bacterium inoculation 4 54.67±3.06 BCc 1.52±0.08 CDd 6.13±0.73 Ed 4.44±0.25 Cc 6 66.67±7.57 ABab 1.69±0.23 Cd 6.77±0.41 DEcd 4.69±0.27 BCbc 对照 Control 31.33±5.03 Dd 0.84±0.11 De 4.48±0.07 Fe 3.93±0.32 Dd 表 5 酸化水不同处理对泰鲜甜2号种子出芽和胚根生长的影响Table 5. Effects of AEOW treatment on seed germination and radicle growth of Taixiantian No. 2组别
Group时间
Time/h出芽率
Germination rate/%芽长
Shoot length/cm主胚根长度
Radicle length/cm胚根数
No of primary root未接种 Without inoculation 4 66.67±7.02 Aab 2.96±0.06 Aab 8.08±0.27 Aa 4.54±0.19 Aa 6 67.33±3.06 Aa 3.00±0.78 Aab 7.84±0.22 Aab 4.25±0.04 ABabc 对照 Control 58.67±3.06 ABabc 2.47±0.19 Aab 7.42±0.14 ABabc 4.18±0.27 ABabcd 真菌接种 Fungus inoculation 4 54.67±5.03 ABc 3.06±0.11 Aa 7.33±0.25 ABbcd 4.07±0.32 ABbcd 6 58.00±6.00 ABabc 2.76±0.20 Aab 7.49±0.52 ABabc 3.97±0.14 ABcd 对照 Control 51.33±4.16 Bc 2.43±0.44 Ab 7.09±0.52 ABcd 3.77±0.31 Bd 细菌接种 Bacterium inoculation 4 56.67±7.02 ABbc 1.45±0.07 Bc 6.63±0.58 Bd 4.42±0.25 Aab 6 61.33±7.57 ABabc 1.43±0.03 Bc 6.78±0.26 Bcd 4.45±0.25 Aab 对照 Control 32.67±4.16 Cd 1.12±0.10 Bc 5.08±0.43 Ce 4.09±0.18 ABbcd 表 6 酸化水处理对玉米出苗和植株生长的影响Table 6. Effects of AEOW on seedling emergence and plant growth of maize品种
Varieties组别
Group出苗率
Seedling emergence rate/%株高
Seedling height/cm地上部鲜重
Fresh weight/g泰鲜甜1号 Taixiantian NO.1 酸化水 AEOW 56.00±2.00 Aa 6.12±0.57 Aa 0.60±0.01 Aa 对照 Control 50.67±3.06 Aa 6.02±0.69 Aa 0.56±0.04 Aa 泰鲜甜2号 Taixiantian NO.2 酸化水 AEOW 62.67±5.03 Aa 6.99±0.95 Aa 0.66±0.12 Aa 对照 Control 54.67±1.15 Aa 6.51±0.77 Aa 0.61±0.03 Aa 2.3 酸化水不同处理对玉米种子真菌和细菌出现频率的影响
在2.2两个品种酸化水不同时间处理的种子中,其细菌、真菌出现频率如表3所示,种子经酸化水处理1 、2 、4 、6 h后,细菌出现的频率分别为15.33%、9.33%、4.67%、1.33%,真菌出现的频率分别为50.00%、32.67%、25.33%和20.00%。表明酸化水处理对种子携带的细菌、真菌有明显的抑制作用,其中以对细菌抑制效果明显优于真菌,随着处理时间越长,酸化水抑制效果越好,种子携带细菌、真菌频率越低。而在携带真菌的种子中,以镰孢菌属真菌出现频率明显高于其他真菌,为种子携带的优势型真菌,如种子经酸化水处理6 h后,镰孢菌属真菌出现频率为16.00%,而曲霉和青霉等真菌的出现频率仅为4.00%。未经酸化水处理的种子多以细菌或以细菌和真菌混合出现,但其真菌种类判别受到了细菌的影响。
2.4 酸化水处理对玉米种子出芽和胚根生长的影响
从表4和表5可知,在未接种外源病原菌的试验组中,与空白对照相比,泰鲜甜1号和泰鲜甜2号种子采用种子与酸化水按(mg/mL)125︰2处理种子4 h和6 h,均能明显提高种子出芽率、促进芽和胚根生长,2个处理时间对种子萌芽的影响效果相当。其中,当种子用酸化水处理6 h后,泰鲜甜1号出芽率、芽长、主胚根长和胚根数分别为73.33%、3.69 cm、9.85 cm和4.92条,比对照分别提高了13.33%、0.32 cm、0.73 cm和0.16条,泰鲜甜2号也比对照分别提高了8.66%、0.55 cm、0.42 cm和0.07条。
而在预先接种外源病原真菌拟轮枝镰孢菌、病原细菌假单孢杆菌组中,与未接种组的空白对照相比,泰鲜甜1号和泰鲜甜2号种子出芽率、芽长、胚根数和主胚根长度明显受到抑制,其中以病原细菌对种子萌发的抑制作用更强,差异达显著性水平。所有接种病菌的种子用酸化水分别处理4 h和6 h后,其出芽率、芽长、胚根数和主胚根长度均有不同程度提高。如在接种病原真菌组中,泰鲜甜1号种子用酸化水分别处理6 h后,其出芽率、芽长、主胚根长度和胚根数分别为68.37%、3.22 cm、9.44 cm和5.16条,分别比其对照(接种病原真菌,未用酸化水处理)提高了17.04%、0.54 cm、1.89 cm和0.60条,而泰鲜甜2号也分别提高了6.67%、0.33 cm、0.40 cm和0.26条。在接种病原细菌组中,泰鲜甜1号种子用酸化水分别处理6 h后,其出芽率、芽长、主胚根长度和胚根数分别为56.67%、1.69 cm、6.78 cm和4.69条,分别比其相应的对照(接种病原细菌,未用酸化水处理)提高35.34%、0.85 cm、2.30 cm和0.76条,而泰鲜甜2号也分别提高了28.66%、0.30 cm、1.70 cm和0.36条,在出芽率方面差异达显著性水平。表明采用酸化水处理玉米种子4~6 h,对种子出芽和生根安全,不同程度上促进了种子萌发和生长,在酸化水对接种2种病菌的种子处理之间,以酸化水对提升细菌接种的种子出芽效果更好。
2.5 酸化水处理对玉米种子出苗和植株生长的影响
从表6可知,供试泰鲜甜1号和泰鲜甜2号其种子与酸化水按(mg/mL)125︰2比例处理6 h后,2个品种出苗率分别为56.00%和62.67%,已出苗植株株高分别为6.12 cm和6.99 cm,地上部鲜重分别为0.60 g和0.66 g,与各品种相应空白对照相比,2个品种在出苗率、株高和地上部鲜重上与空白对照无显著性差别,但出苗率分别比对照提高了5.33%和8.00%,不同程度上提高了苗的株高和地上部鲜重。表明酸化水处理对玉米种子出苗和植株生长安全,能明显提高种子出苗率,促进苗的生长。
3. 讨论与结论
酸化水是一种具有高氧化还原电位(ORP),低pH值,含低浓度活性氯、活性氧和次氯酸的水,属非化学品种制剂,其独特的杀菌作用机理,能作用多种病原微生物,被认为是一种环保、安全、无残留的广谱性新型杀菌消毒剂。本研究将玉米种子表面微生物的菌悬液、玉米种子与酸化水分别按不同比例混匀后,发现酸化水处理对玉米种子表面微生物有抑制作用,且随着酸化水用量增加及作用时间的延长,其抑菌效果越好,从菌悬液中和种子中检测出的菌量越少,种子带菌率越低。泰鲜甜1号、泰鲜甜2号种子菌悬液与酸化水按(mL/mL)1︰2比例及种子与酸化水按(mg/mL)125︰2比例混合6 h的处理便可达较好的抑菌效果。
高新昊等[26]的研究表明采用适宜有效氯浓度电解水浸种可提高种子胚芽生长速率,促进黄瓜幼苗生长与干物质积累,提高黄瓜幼苗的质量。Liu等[27]采用酸化水浸种可促进糙米芽生长,提高其γ-氨基丁酸含量。赵峰等[28]研究发现,酸性氧化电解水浸泡对水稻种子发芽及生长有促进作用,与清水对照相比,能增加根长、芽长和苗鲜重,而用咪鲜胺浸泡水稻种子对苗期芽的生长有抑制作用。本研究结果表明,采用酸化水浸泡处理玉米种子4 h和6 h,对种子出芽和生根安全,与清水对照相比,能明显提高种子出芽率、促进芽和胚根生长,用酸化水浸泡处理玉米种子6 h,对种子出苗安全,能明显提高种子出苗率,促进苗的生长,增加鲜重。
由于不清楚本研究供试玉米种子携带的微生物是否为致病菌,同时为验证种子携带病原菌及酸化水对种子出芽现状的影响,本研究采用分离自玉米鞘腐病的病原真菌(拟轮枝孢菌)和玉米茎腐病的病原细菌(假单胞杆菌)分别进行泰鲜甜1号和泰鲜甜2号玉米种子人工接菌处理,试验结果表明,预先接种的2种病原菌均能影响种子出芽和胚根的生长,而采用酸化水浸泡处理病种4 h和6 h,能明显改善种子的出芽性状,提高出芽率,促进胚根生长,以酸化水处理种子6 h的效果优于4 h。表明酸化水处理可能通过抑制玉米种子外源病原菌的生长,从而减少病原菌对种子出芽和生根的负面影响。基于上述研究结果表明,采用酸化水浸泡处理玉米种子6 h进行种子表面微生物或病原菌的消毒处理是可行的,但在增加作用时间条件下,酸化水对种子出芽、出苗的影响是否安全,以及酸化水浸种处理是否能通过影响种子的生理生化特性,促进出苗和提高抗逆性,有待进一步研究。
黄思良等[29]从玉米穗腐病病粒上分离出多种致病性镰孢菌,而本研究也证实,玉米种子表面携带有大量微生物,包括细菌、真菌等,其中镰孢菌在真菌中检出频率最高,因此,做好种子的杀菌处理是预防后期病害发生的重要环节。用酸化水处理种子用量少,酸化水的使用可减轻因种子表面病原菌原因导致的烂种,对促进种子出芽和出苗生长,降低种传病害发生的初侵染源具有重要作用。但酸化水不同条件下贮存,对其理化性能和杀菌效果均有影响,如54 ℃存放14 d条件下会出现氧化还原电位下降、pH值升高[30],在常温、封闭、不避光的保存条件下,有效期为7-10 d[31]。因此,应在适宜条件下保存酸化水,有条件时最好是现制现用。另从酸化水处理后的种子携带细菌、真菌出现频率结果分析,酸化水处理对种子携带的细菌抑制效果明显优于真菌,在酸化水对预先接种两类病原菌的种子处理之间,也以酸化水对提升接种细菌种子的出芽性状更好,推测酸化水对由病原细菌引起的种传病害可能具有更好的应用前景。本研究结果可为其他作物种子病原菌的杀菌处理提供参考,同时为作物生产过程中预防健田病害发生的风险性、阻隔种传病害途径发挥重要作用,也为化学农药减施增效途径提供参考。
-
![]()
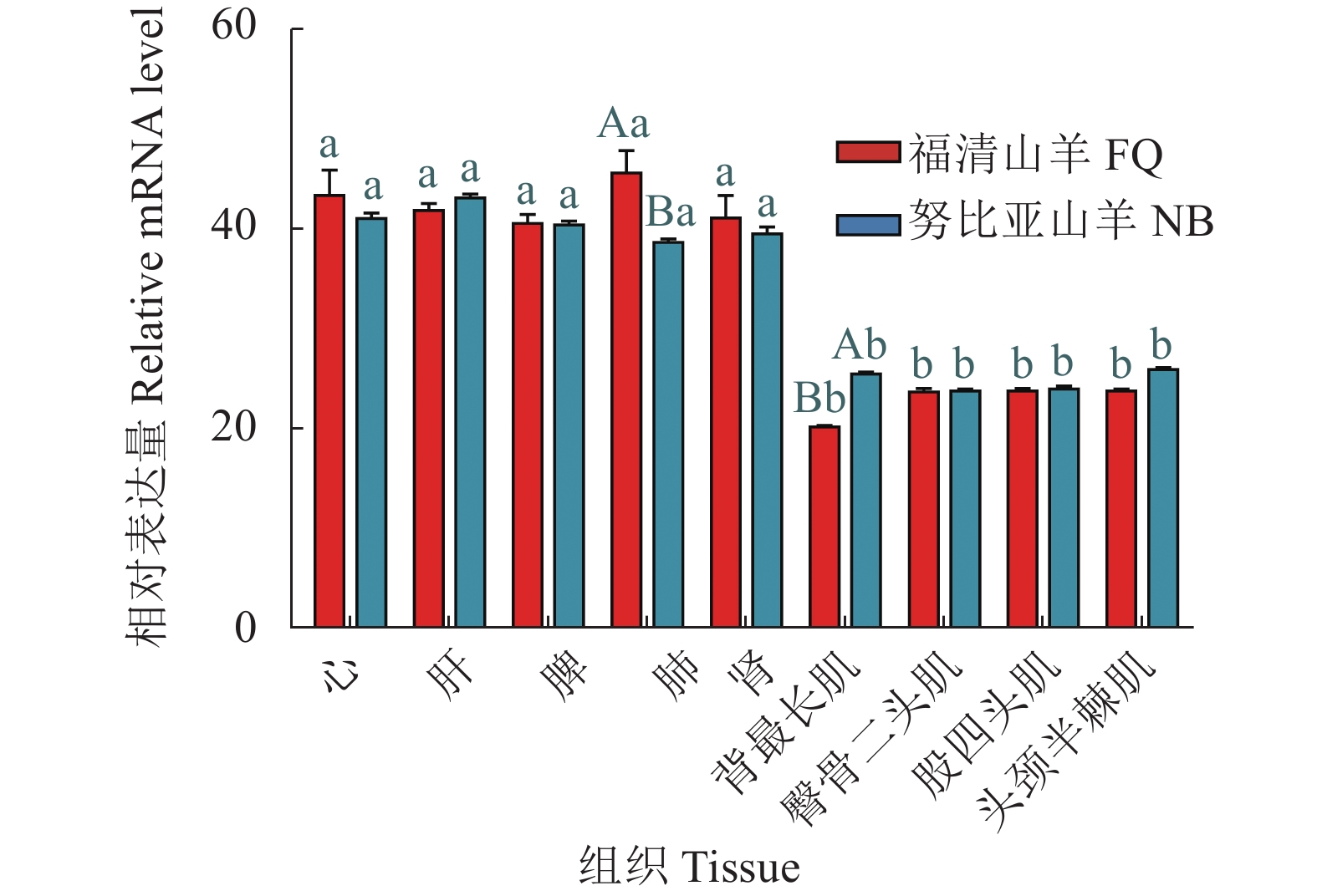
图 1 IL15RA基因在福清山羊与努比亚山羊各组织的表达量
不同小写字母表示同一山羊品种不同组织之间差异显著(P<0.05);不同大写字母表示福清山羊和努比亚山羊同一组织间差异显著(P<0.05)。
Figure 1. Expressions of IL15RA in tissues of Fuqing and Nubian goats
Different lowercase letters represent significantly differences between different tissues of Fuqing and Nubian goats (P<0.05); different uppercase letters, significant differences on same tissue between Fuqing and Nubian goats (P<0.05).
![]()
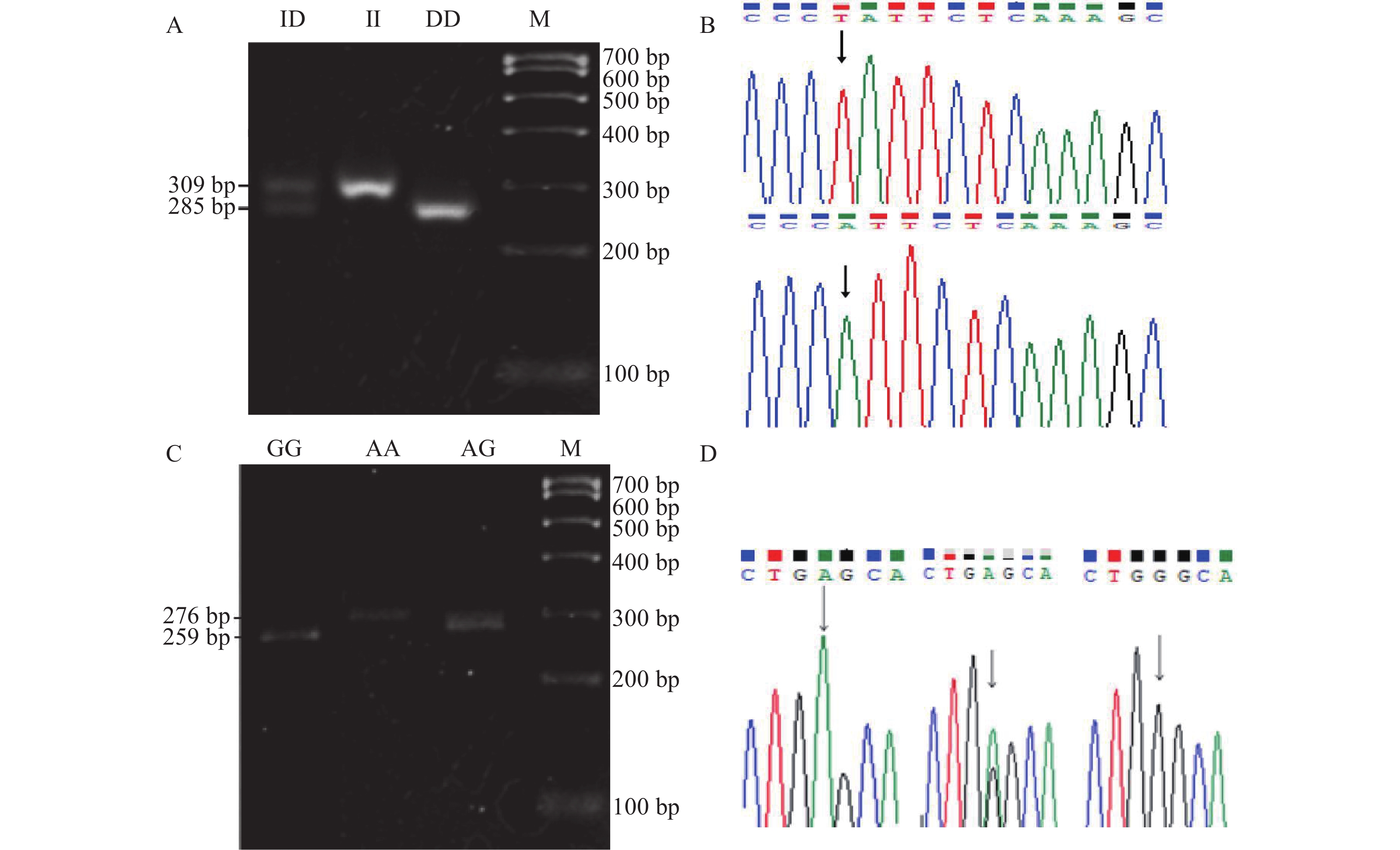
图 2 山羊IL15RA基因位点检测结果
M为MakerⅠ,图A、B为Del22224、图C、D为g33428A>G的RFLP分型电泳图谱及测序峰图。
Figure 2. Localization of IL15RA
M: Maker Ⅰ, A and B: RFLP typing electrophoretic profiles and sequencing peaks of Del22224; C and D: RFLP typing electrophoretic profiles and sequencing peaks of g33428A>G.
表 1 引物信息
Table 1 Information on primers
引物
Primer引物序列(5′-3′)
Sequence of primer (5′-3′)退火温度
Temperature/
℃产物大小
Product
size/bp用途
PurposesP1 F:ACTGACTTCCACCCAA
GCATC60 780 筛选突变位点 R:CTCAGGGCAGGTATC
AACAGGP2 F:TGCTGTGTGGTTATCC
TCTCT58 573 筛选突变位点 R:CTTAGGCCTACTTAA
GGCACGP3 F:GACCATGGCAATAACT
GAGCTG54 487 筛选突变位点 R:GCCTTTTATACCAAGA
ACGCGAP4 F:TGACACAATCTCTCCT
CTGTCA54 838 筛选突变位点 R:TACAAGGAACAGTTT
CAAAATACGAP5 F:CCCCTCCGGAAAAGGT
AGGA60 856 InDel R:GAGGGTGTCAGAAGA
CACAGGP6 F:TGCCGACTCATTCTGC
TCTC60 588 SNP R:TGAAATGTCAGCAAG
TGGCAGP7 F:CCACCCACTCTGTGC
CG63 276 PCR-RFLP
分型R:TATGGGGACGTGTGTC
CATCTP8 F:TGGTAGATGCGTAGCT
TTGAGGAT62 309 PCR-RFLP
分型R:GAGCACAGGGCAGAT
GTGTTCAP9 F:AAGGAAGACCCTTTCC
TGGCT61 149 qRT-PCR R:TGCCTGGATTCTGACC
AGTGAP10 F:GCAAGGACCTTTACGC
CAAC59 116 内参基因 R:CTTGATCTTCATCGTG
CTGGG 下载: 导出CSV
下载: 导出CSV
表 2 山羊IL15RA基因突变位点基因型频率和等位基因频率
Table 2 Genotypic and allelic frequencies on polymorphism of IL15RA mutation
位点
Locus品种
Breeds基因型频率
Genotypic frequencies/%基因频率
Allelic frequencies/%有效等位基因数
Ne杂合性
He多态信息含量
PIC哈迪-温伯格检验
X2II ID DD I D Del22224 福清山羊 (n=108) 0.385 0.519 0.096 0.644 0.356 1.846 0.458 0.353 P<0.05 努比亚山羊(n=286) 0.688 0.271 0.041 0.823 0.177 1.410 0.291 0.249 P<0.05 SNP-
g.33428AA AG GG A G 福清山羊 (n=108) 0.769 0.193 0.038 0.866 0.134 1.303 0.232 0.205 P<0.05 努比亚山羊 (n=286) 0.280 0.360 0.360 0.460 0.540 1.987 0.497 0.373 P>0.05
下载: 导出CSV
表 3 山羊IL15RA基因突变位点与山羊生长性状的关联分析
Table 3 Relationship between IL15RA mutation site and growth of goat
品种
Breeds生长性状
Growth traitsDel22224位点基因型
Genotypes of Del22224SNP-g.33428位点基因型
Genotypes of SNP-g.33428II ID DD AA AG GG 福清山羊FQ 体重/kg 30.84±1.05 29.50±0.90 30.18±2.02 30.74±0.69a 33.93±3.94a 26.51±1.31b 体高/cm 55.29±0.66 54.54±0.49 55.49±1.22 55.04±0.37 57.10±1.41 53.85±1.13 体长/cm 55.27±0.74 53.97±0.54 54.84±2.22 55.07±0.52 55.50±1.62 53.83±1.12 胸围/cm 73.35±1.08 72.89±0.88 71.92±1.98 73.82±0.64a 77.63±3.77a 68.78±1.55b 胸宽/cm 16.25±0.27 16.37±0.25 16.76±0.68 16.68±0.18a 16.60±0.81ab 15.17±0.44b 胸深/cm 29.45±0.58 30.02±0.39 29.06±0.93 30.19±0.33a 30.70±1.00a 27.78±0.71b 管围/cm 7.48±0.10 7.45±0.08 7.40±0.23 7.47±0.07ab 7.95±0.18a 7.23±0.14b 尻宽/cm 14.83±0.24 14.97±0.21 15.00±0.35 15.22±0.14a 15.00±0.82ab 13.68±0.33b 体躯指数 131.91±1.87 135.17±1.19 132.62±5.58 134.54±1.22a 139.68±2.76a 127.99±2.08b 体长指数 101.19±1.46 99.24±1.17 98.96±3.58 100.36±1.08 97.23±2.21 100.23±1.62 胸围指数 132.87±1.71 133.92±1.65 129.87±3.63 134.44±1.28a 135.93±5.64a 127.94±2.05b 管围指数 13.57±0.22 13.68±0.14 13.35±0.34 13.60±0.12 13.93±0.25 13.50±0.30 胸宽指数 56.07±1.71 54.64±0.66 57.65±1.45 55.72±0.89 54.03±1.33 54.77±1.25 髋骨指数 110.18±1.88 109.66±1.30 111.63±3.36 109.97±1.20 110.80±1.11 111.03±2.00 努比亚山羊NB 体重/kg 47.03±0.77 48.50±1.23 49.26±3.68 47.68±1.23 48.51±1.07 47.26±1.04 体高/cm 68.45±0.42 69.50±0.74 67.75±1.51 67.86±0.61 69.10±0.61 69.23±0.56 体长/cm 64.78±0.38 65.26±0.65 66.80±1.29 64.96±0.58 65.50±0.51 64.62±0.52 胸围/cm 86.08±0.54 86.67±0.85 87.91±2.44 86.39±0.79 86.73±0.75 86.16±0.74 胸宽/cm 20.31±0.23 20.20±0.40 21.13±1.01 19.82±0.29 20.50±0.33 20.66±0.34 胸深/cm 33.61±0.29 34.08±0.38 34.46±1.07 33.68±0.39 34.12±0.37 33.55±0.38 管围/cm 10.25±0.46 10.13±0.17 10.73±0.28 9.83±0.12 10.82±0.82 9.79±0.10 尻宽/cm 18.06±0.14 18.27±0.27 18.75±0.69 17.99±0.18 18.34±0.22 17.98±0.20 体躯指数 133.18±0.72 133.21±1.14 131.64±2.74 133.35±1.13 132.74±1.01 133.57±0.86 体长指数 94.95±0.55 94.25±0.89 99.03±2.62 96.08±0.91a 95.14±0.72ab 93.62±0.71b 胸围指数 126.07±0.69 125.17±1.14 130.06±3.59 127.52±0.86 125.94±1.00 124.81±1.00 管围指数 15.03±0.66 14.64±0.25 15.91±0.53 14.53±0.18 15.73±1.18 14.21±0.16 胸宽指数 60.60±0.53 59.22±0.89 61.10±1.46 58.96±0.65b 60.08±0.69ab 61.70±0.77a 髋骨指数 113.09±1.23 111.11±1.88 112.96±4.46 110.62±1.58 112.44±1.66 115.48±1.79 不同字母表示同一山羊品种同一位点不同基因型之间差异显著(P<0.05),未标注字母表示差异不显著(P≥0.05)。表4、5同。
Data with different letters indicate significant differences between genotypes of the same mutation site of Fuqing or Nubian goat. (P<0.05), no letters, no significant differences(P≥0.05). Same for Table 4, 5.
下载: 导出CSV
表 4 福清山羊IL15RA基因单倍型组合与生长性状关联分析
Table 4 Correlation between IL15RA haplotype combination and growth traits of Fuqing goat
生长性状
Growth traits单倍型组合 Haplotype combination DD/AA DD/AG ID/AA ID/AG ID/GG II/AA II/AG 体重 Body weight/kg 32.03±1.06a 24.60±2.95b 29.69±1.01ab 26.70±2.03b 33.93±3.94a 30.17±3.15ab 30.20±2.43ab 体高 Body height/cm 55.66±0.55 52.72±3.20 54.49±0.52 53.63±1.64 57.10±1.41 55.25±2.03 55.85±0.92 体长 Body length/cm 56.19±0.71 53.53±3.14 54.03±0.66 53.01±1.02 55.50±1.62 53.20±3.27 56.30±2.64 胸围 Chest circumference/cm 74.72±0.98a 65.83±3.66b 72.93±0.96a 70.59±2.34ab 77.63±3.77a 73.72±2.46a 69.23±3.19ab 胸宽 Chest width/cm 30.07±0.61a 25.98±1.16b 30.17±0.42a 29.01±1.32ab 30.70±1.00a 30.35±1.22a 27.13±0.85ab 胸深 Chest depth/cm 16.55±0.26a 14.50±0.84b 16.61±0.27a 15.18±0.70ab 16.60±0.81a 17.43±0.75a 15.75±1.22ab 管围 Cannon circumference/cm 7.52±0.11 7.22±0.34 7.44±0.09 7.24±0.22 7.95±0.18 7.45±0.33 7.33±0.33 尻宽 Hucklebone width/cm 15.15±0.22a 12.85±0.60b 15.23±0.22a 14.59±0.57ab 15.00±0.82a 15.33±0.44a 14.50±0.56ab 体躯指数 Trunk index 133.45±2.05a 123.45±4.28b 135.23±1.46a 132.91±2.14ab 139.68±2.76a 140.28±7.26a 121.13±5.31b 体长指数 Body length index 101.26±1.64 101.92±3.82 99.44±1.47 99.22±2.00 97.23±2.21 96.52±5.19 102.63±4.60 胸围指数 Chest circumference index 134.49±1.84 125.35±4.01 134.19±2.01 131.81±3.19 135.93±5.64 133.90±4.86 123.83±4.43 管围指数 Cannon circumference index 13.53±0.21 13.87±0.95 13.68±0.17 13.54±0.30 13.93±0.25 13.52±0.50 13.10±0.45 胸宽指数 Chest width index 56.03±2.01 56.08±3.11 55.20±0.80 52.38±1.26 54.03±1.33 57.48±1.20 57.90±3.49 髋骨指数 Huckle bone width index 109.73±2.10 113.15±5.09 109.43±1.61 110.18±2.76 110.80±1.11 113.97±4.75 108.13±4.63
下载: 导出CSV
表 5 努比亚山羊IL15RA 基因单倍型组合与生长性状关联分析
Table 5 Correlation between IL15RA haplotype combination and growth traits of Nubian goat
生长性状
Growth traits单倍型组合 Haplotype combination DD/AA DD/AG DD/GG ID/AA ID/AG ID/GG II/AG II/GG 体重
Body weight/kg47.77±1.54ab 45.60±1.24b 47.67±1.37ab 52.26±2.10a 46.97±1.80ab 49.42±1.81ab 48.64±2.66ab 55.09±5.80a 体高
Body height/cm67.62±0.75b 69.10±0.71ab 68.61±0.68ab 69.68±0.82ab 69.51±1.15ab 70.53±1.09a 69.79±1.44ab 72.25±1.45a 体长
Body length/cm64.72±0.67 64.61±0.64 65.38±0.65 67.14±1.21 65.24±0.67 65.26±1.09 67.63±0.98 67.65±1.72 胸围
Chest circumference/
cm86.45±0.97 85.86±0.95 86.31±0.86 88.79±1.14 84.98±1.01 87.98±1.53 89.89±2.54 88.28±3.36 胸宽
Chest width/cm19.97±0.32 20.97±0.43 20.61±0.42 20.37±0.55 20.20±0.65 21.34±0.63 21.10±0.93 21.40±1.19 胸深
Chest depth/cm33.70±0.44 33.44±0.50 34.26±0.49 34.61±0.60 34.40±0.56 34.31±0.71 34.59±0.88 35.32±1.70 管围
Cannon circumference/
cm10.01±0.16 9.81±0.14 11.50±1.28 10.49±0.25 10.14±0.20 10.49±0.38 10.67±0.19 10.73±0.59 尻宽
Hucklebone width/cm18.00±0.26b 18.04±0.23b 18.27±0.29ab 19.23±0.42a 18.47±0.31ab 18.94±0.46a 18.31±0.41ab 19.27±0.48a 体躯指数
Trunk index133.90±1.28 133.09±1.09 132.40±1.29 132.66±2.32 130.48±1.65 135.17±1.83 132.94±3.51 130.50±3.58 体长指数
Body length index96.10±1.01a 93.82±0.90ab 95.55±0.89ab 96.49±1.97a 94.36±1.45ab 92.74±1.31b 97.23±2.78a 93.70±2.12ab 胸围指数
Chest circumference
index128.18±1.20a 124.54±1.14b 126.17±1.21ab 127.49±1.31ab 122.83±1.90b 124.94±1.58ab 129.00±3.71a 122.08±3.10b 管围指数
Cannon circumference
index14.89±0.26ab 14.28±0.23b 16.83±1.85a 15.09±0.43ab 14.70±0.40ab 14.93±0.54ab 15.30±0.30ab 14.83±0.72ab 胸宽指数
Chest width index59.43±0.79b 62.81±0.89a 60.17±0.87ab 58.99±1.62b 58.64±1.46b 62.24±1.30ab 61.03±2.18ab 60.68±2.08ab 髋骨指数
Huckle bone width
index111.68±1.78ab 116.73±2.22a 113.53±2.17ab 106.54±3.56b 109.79±3.57ab 113.35±2.81ab 115.39±4.89ab 111.33±6.56ab
下载: 导出CSV
-
[1] 陈甜, 肖海峰. 中国羊肉消费状况及影响因素研究 [J]. 中国畜牧杂志, 2016, 52(12):15−20. CHEN T, XIAO H F. Research on the factors and the influence of mutton consumption situation in China [J]. Chinese Journal of Animal Science, 2016, 52(12): 15−20. (in Chinese)
[2] 刘远, 李文杨, 吴贤锋, 等. 福清山羊与努比亚黑山羊背最长肌比较转录组分析 [J]. 中国农业科学, 2019, 52(14):2525−2537. DOI: 10.3864/j.issn.0578-1752.2019.14.011 LIU Y, LI W Y, WU X F, et al. Transcriptome analysis of differentially gene expression associated with longissimus Doris tissue in Fuqing goat and Nubian black goat [J]. Scientia Agricultura Sinica, 2019, 52(14): 2525−2537. (in Chinese) DOI: 10.3864/j.issn.0578-1752.2019.14.011
[3] MORRIS R, KERSHAW N J, BABON J J. The molecular details of cytokine signaling via the JAK/STAT pathway [J]. Protein Science, 2018, 27(12): 1984−2009. DOI: 10.1002/pro.3519
[4] RICHARD A J, STEPHENS J M. The role of JAK–STAT signaling in adipose tissue function [J]. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease, 2014, 1842(3): 431−439. DOI: 10.1016/j.bbadis.2013.05.030
[5] FUJIMOTO M, NAKA T. Regulation of cytokine signaling by SOCS family molecules [J]. Trends in Immunology, 2003, 24(12): 659−666. DOI: 10.1016/j.it.2003.10.008
[6] HEWETSON A, MOORE S L, CHILTON B S. Prolactin signals through RUSH/SMARCA3 in the absence of a physical association with Stat5a [J]. Biology of Reproduction, 2004, 71(6): 1907−1912. DOI: 10.1095/biolreprod.104.031435
[7] LEONARD W J, O’SHEA J J. Jaks and STATs: Biological implications [J]. Annual Review of Immunology, 1998, 16: 293−322. DOI: 10.1146/annurev.immunol.16.1.293
[8] ZHU J, SHIMIZU E, ZHANG X R, et al. EGFR signaling suppresses osteoblast differentiation and inhibits expression of master osteoblastic transcription factors Runx2 and Osterix [J]. Journal of Cellular Biochemistry, 2011, 112(7): 1749−1760. DOI: 10.1002/jcb.23094
[9] GABER T, BRINKMAN A C K, PIENCZIKOWSKI J, et al. Impact of Janus kinase inhibition with tofacitinib on fundamental processes of bone healing [J]. International Journal of Molecular Sciences, 2020, 21(3): 865. DOI: 10.3390/ijms21030865
[10] AKAMATSU N, NAKAJIMA H, OHNO M, et al. Increase in acetyl CoA synthetase activity after phenobarbital treatment [J]. Biochemical Pharmacology, 1975, 24(18): 1725−1727. DOI: 10.1016/0006-2952(75)90013-1
[11] GRABSTEIN K H, EISENMAN J, SHANEBECK K, et al. Cloning of a T cell growth factor that interacts with the beta chain of the interleukin-2 receptor [J]. Science, 1994, 264(5161): 965−968. DOI: 10.1126/science.8178155
[12] GILBERT R P, BAILEY D R, SHANNON N H. Linear body measurements of cattle before and after 20 years of selection for postweaning gain when fed two different diets [J]. Journal of Animal Science, 1993, 71(7): 1712−1720. DOI: 10.2527/1993.7171712x
[13] LI W Y, LIU Y, GAO C F, et al. A novel duplicated insertion/deletion (InDel) of the CPT1a gene and its effects on growth traits in goat [J]. Animal Biotechnology, 2021, 32(3): 343−351. DOI: 10.1080/10495398.2019.1698433
[14] 吴贤锋, 王金宝, 刘远, 等. 基于转录组测序的ITGAD基因InDel位点鉴定及其与山羊生长性状的关联性分析 [J]. 农业生物技术学报, 2022, 30(9):1763−1770. DOI: 10.3969/j.issn.1674-7968.2022.09.010 WU X F, WANG J B, LIU Y, et al. Identification of InDel locus of ITGAD gene based on transcriptome sequencing and its association with goat (Capra hircus) growth traits [J]. Journal of Agricultural Biotechnology, 2022, 30(9): 1763−1770. (in Chinese) DOI: 10.3969/j.issn.1674-7968.2022.09.010
[15] WU X F, LIU Y, ZHAN J S, et al. A novel splice variant of goat CPT1a gene and their diverse mRNA expression profiles [J]. Animal Biotechnology, 2023, 34(7): 2571−2581. DOI: 10.1080/10495398.2022.2106573
[16] KLINKENBERG E, ONWONA-AGYEMAN K A, MCCALL P J, et al. Cohort trial reveals community impact of insecticide-treated nets on malariometric indices in urban Ghana [J]. Transactions of the Royal Society of Tropical Medicine and Hygiene, 2010, 104(7): 496−503. DOI: 10.1016/j.trstmh.2010.03.004
[17] WU X F, LIU Y, WANG Y G, et al. A novel 22-bp InDel within FGF7 gene is significantly associated with growth traits in goat [J]. Animal Biotechnology, 2024, 35(1): 2262537. DOI: 10.1080/10495398.2023.2262537
[18] SVINKA J, MIKULITS W, EFERL R. STAT3 in hepatocellular carcinoma: New perspectives [J]. Hepatic Oncology, 2014, 1(1): 107−120. DOI: 10.2217/hep.13.7
[19] MOHRI T, IWAKURA T, NAKAYAMA H, et al. JAK-STAT signaling in cardiomyogenesis of cardiac stem cells [J]. JAK-STAT, 2012, 1(2): 125−130. DOI: 10.4161/jkst.20296
[20] LI J L. JAK-STAT and bone metabolism [J]. Jak-Stat, 2013, 2(3): e23930.
[21] CORRY K A, ZHOU H K, BRUSTOVETSKY T, et al. Stat3 in osteocytes mediates osteogenic response to loading [J]. Bone Reports, 2019, 11: 100218.
[22] PISTILLI E E, BOGDANOVICH S, GARTON F, et al. Loss of IL-15 receptor α alters the endurance, fatigability, and metabolic characteristics of mouse fast skeletal muscles [J]. Journal of Clinical Investigation, 2011, 121(8): 3120−3132.
[23] LORO E, RAMASWAMY G, CHANDRA A, et al. IL15RA is required for osteoblast function and bone mineralization [J]. Bone, 2017, 103: 20−30.
[24] LORO E, BISETTO S, KHURANA T S. Mitochondrial ultrastructural adaptations in fast muscles of mice lacking IL15RA [J]. Journal of Cell Science, 2018, 131(21): jcs218313.
[25] KORNSUTHISOPON C, MANOKAWINCHOKE J, SONPOUNG O, et al. Interleukin 15 participates in Jagged1-induced mineralization in human dental pulp cells [J]. Archives of Oral Biology, 2021, 128: 105163.
[26] LIU Y, WU X F, XU Q, et al. Temporal transcriptome dynamics of Longissimus dorsi reveals the mechanism of the differences in muscle development and IMF deposition between Fuqing goats and Nubian goats [J]. Animals, 2024, 14(12): 1770.
[27] 刘远, 李文杨, 吴贤锋, 等. 福清山羊快长品系的世代选育及生产性能测定分析 [J]. 福建农业学报, 2021, 36(3):305−311. LIU Y, LI W Y, WU X F, et al. Growth and reproduction of newly bred fast-growing Fuqing goats in two generations [J]. Fujian Journal of Agricultural Sciences, 2021, 36(3): 305−311. (in Chinese)
[28] 曹艳红. 利用高通量测序解析隆林山羊与努比亚山羊的遗传差异及表达差异[D]. 杨凌: 西北农林科技大学, 2019. CAO Y H. Analysis of genetic differences and expression differences between Longlin goat and Nubian goat by high-throughput sequencing[D]. Yangling: Northwest A & F University, 2019. (in Chinese)
[29] MAQUAT L E. Nonsense-mediated mRNA decay: Splicing, translation and mRNP dynamics [J]. Nature Reviews Molecular Cell Biology, 2004, 5(2): 89−99. DOI: 10.1038/nrm1310
[30] LU S H, CULLEN B R. Analysis of the stimulatory effect of splicing on mRNA production and utilization in mammalian cells [J]. RNA, 2003, 9(5): 618−630. DOI: 10.1261/rna.5260303
[31] NOTT A, MEISLIN S H, MOORE M J. A quantitative analysis of intron effects on mammalian gene expression [J]. RNA, 2003, 9(5): 607−617. DOI: 10.1261/rna.5250403
[32] 刘玉, 张林林, 房义, 等. 湖羊STAT5a基因第10内含子多态性及其与泌乳性状的关联分析 [J]. 中国畜牧兽医, 2023, 50(9):3680−3687. LIU Y, ZHANG L L, FANG Y, et al. Polymorphism of STAT5a gene intron 10 and its association with lactation traits in hu sheep [J]. China Animal Husbandry & Veterinary Medicine, 2023, 50(9): 3680−3687. (in Chinese)
[33] 李志斌. 简州大耳羊ENO3和TBC1D7基因多态性及其与生长性状的关联分析[D]. 成都:西南民族大学, 2023. LI Z B. Association analysis of ENO3 and TBC1D7 genetic polymorphisms with growth traits in Jianzhou Da'er goat[D]. Chengdu: Southwest Minzu University, 2023. (in Chinese)
[34] ZHOU S L, SHI X Y, SONG C C, et al. SNP discovery of PRKAB1 gene and their associations with growth traits in goats [J]. Animal Biotechnology, 2022, 33(7): 1613−1619. DOI: 10.1080/10495398.2021.1920426
[35] 阮涌, 陈祥, 田贵刚, 等. F3代波杂山羊XKR4 基因多态性与生长性状的关联分析 [J]. 南方农业学报, 2022, 53(6):1752−1758. RUAN Y, CHEN X, TIAN G G, et al. Correlation analysis of XKR4 gene polymorphism and growth traits in F3 generation Boza goats [J]. Journal of Southern Agriculture, 2022, 53(6): 1752−1758. (in Chinese)
[36] 杨韩, 张阳海, 王敏, 等. 陕北白绒山羊POU1F1基因3’-UTR多态性及其与生长性状的相关分析 [J]. 农业生物技术学报, 2019, 27(7):1224−1232. YANG H, ZHANG Y H, WANG M, et al. Polymorphisms of 3'-UTR of POU1F1 gene and its association with growth traits in shaanbei white Cashmere goats(Capra hircus) [J]. Journal of Agricultural Biotechnology, 2019, 27(7): 1224−1232. (in Chinese)
计量
- 文章访问数: 64
- HTML全文浏览量: 17
- PDF下载量: 18
