

Study on Transcriptome of Moringa oleifera Lam. Leaves Based on RNA-seq
-
摘要: 为探讨辣木叶营养价值形成的遗传基础,采用RNA-seq对不同发育时期的辣木叶混合样品进行转录组测序,获得44 056 449个reads,共组装得到75 452条unigene,平均长度为795.69 nt,其中有15 824条unigene长度大于1 000 nt。采用公共数据库进行同源比对,共有30 847条unigene得到注释。其中,有18 277条unigenes被注释到GO数据库的52个功能组,9 689条unigene在COG数据库中得到注释。有10 130条unigene被富集到129条KEGG途径,包括碳代谢、氨基酸生物合成和氧化磷酸化途径。Abstract: To explore the genetic basis for the nutritive value of Moringa oleifera leaves, the leaves at different developmental stages were mixed and used for transcriptome sequencing in the present study. With RNA-seq, 44 056 449 reads were generated, then assembled into 75 452 unigenes with an average length of 795.69 nt. 15 824 of these unigenes were longer than 1 kb. 30 847 unigenes were annotated by a similarity search against public databases. Of these, 18 277 unigenes were assigned into 52 GO function groups and 9 689 unigenes were annotated by COG database. 10 130 unigenes were enriched into 129 KEGG pathways, including carbon metabolism, biosynthesis of amino acids and oxidative phosphorylation.
-
Keywords:
- Moringa oleifera /
- transcriptome /
- function annotation
-
辣木Moringa oleifera Lam.是一种原产亚洲的热带树种,在亚洲、非洲和美洲的热带和亚热带地区均有栽培[1]。辣木各个部分均可食用或作为药物用于治疗多种炎症引发的疾病[2],其中辣木叶因具有很高的营养价值和药用价值而备受关注[3-4]。辣木叶富含蛋白质、矿物质、β-胡萝卜素和维生素多种营养物质[5]。辣木叶的蛋白质含量高于牛奶[6],并含有所有必需氨基酸[7]。研究表明辣木叶的维生素A含量高于胡萝卜,维生素C含量高于柑橘,钙含量高于牛奶,铁含量高于菠菜,钾含量高于香蕉[8]。6勺辣木叶粉即可满足孕妇和哺乳期妇女对铁和钙日需求量[7]。辣木叶的提取物具有抗菌、抗炎症和抗癌等活性[9-11],是许多国家用于治疗多种疾病的传统药物[4]。此外,辣木叶还是优质的蛋白质饲料[12],具有十分广阔的开发利用前景。
目前,关于辣木的研究主集中在营养、功能成分鉴定、食品开发利用等方面[6, 9, 13-14]。仅有少量关于辣木营养和药用品质形成分子基础的报道。Tian等[15]完成了辣木基因组的测序,并在此基础上对辣木转录因子进行了预测与分析[16]。Kondo等[17]从辣木叶中分离得到了抗坏血酸合成酶基因,并对其表达模式进行了分析,发现该基因在叶片中表达量较高。Deng等[18]筛选出2个用于辣木实时荧光定量PCR分析的内参基因,并进一步筛选了辣木黄酮类化合物、萜类化合物和脂肪酸合成相关基因[19]。但辣木大量功能基因的注释亟待完善。本研究对不同发育阶段辣木叶片混合样品进行转录组测序和组装,并对组装得到的unigene进行功能注释,以期为辣木叶营养和药用品质相关功能基因挖掘及其分子机制解析奠定基础。
1. 材料与方法
1.1 试验材料
本试验所用材料为3年生印度辣木PKM2不同发育阶段的叶片(展叶期后0、10、20、30 d),样品取自福建省农业科学院果树研究所辣木资源圃。于同一棵树上摘取生长良好无病虫害的辣木叶片,采下后去除叶柄,迅速用液氮速冻后保存于-80℃冰箱。
1.2 RNA提取、文库构建和RNA-seq
采用EZNA Plant RNA Kit (Omega Bio-tek)进行不同阶段辣木叶片总RNA的提取。采用Qubit 2.0 Fluorometer (Invitrogen, Life Technologies, CA, USA)和Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA)进行RNA浓度的测定和质量评价。将4个阶段辣木叶片的RNA等量混合后,用于测序文库构建。辣木叶片RNA提取、文库构建和RNA-seq由北京百迈客生物科技有限公司完成。cDNA文库的构建参照Fang等[20]的方法。采用Illumina HiSeq 4000高通量测序平台对cDNA文库进行测序。测序原始数据已上传至NCBI,登录号为SRR5765086。
1.3 转录组的组装与表达分析
对Raw reads进行数据过滤,去除其中的接头序列及带有N或10%的碱基Q值≤20的低质量reads,获得高质量的Clean Data。获得高质量的测序数据之后,采用Trinity法对其进行序列组装[21]。所有参数均为默认参数。首先将测序reads打断为较短的片段(K-mer),然后将这些小片段延伸成较长的片段(Contig),并利用这些片段之间的重叠,得到片段集合,最后利用De Bruijn图的方法和测序Read信息,在各个片段集合中分别识别转录本序列。采用Bowtie将各样品测序得到的reads与unigene库进行比对[22],根据比对结果,结合RSEM进行表达量水平估计[23]。利用FPKM值表示对应unigene的表达丰度。
1.4 功能注释和分类
采用BLASTx进行辣木叶片unigene序列的注释(E值<10-5),所用的数据库为Nr、UniProt/Swiss-Prot、GO、COG、KEGG、KOG、Pfam和eggNOG。采用Blast2GO[24]对unigene进行GO注释(E值≤10-5)。GO功能分类结果采用WEGO[25]进行作图。
2. 结果与分析
2.1 辣木叶RNA-Seq测序与组装
为了获得尽可能丰富的辣木叶转录组信息,选取展叶期及展叶期后10、20和30 d的辣木叶片,分别提取不同时期辣木叶片的总RNA,并等量混合,用于构建文库和RNA-Seq测序。HiSeq4000测序和数据过滤后,共获得44 056 449个reads,总长度为13 082 788 218(13.08 Gb)个碱基,GC含量为46.32%。碱基质量值Q30为93.95%。
采用Trinity软件进行辣木叶转录组组装,共得到223 983条转录本,平均长度为1 957.79 nt(表 1)。共得到75 452条unigene,总长度60 036 488 nt,平均长度为795.69 nt,N50长度为1 543 nt,组装完整性较高(表 1)。其中,长度为200~500 nt的unigene有45 964条(60.92%),长度为500~1 000 nt的unigene有13 664条(18.11%),长度大于1 000 nt的unigene有15 824条(20.98%)。
表 1 辣木叶转录组组装结果Table 1. Summary of sequencing and de novo assembly of Moringa oleifera Lam. leaves长度区间/nt 转录本/条 unigenes/条 200~300 30299(13.53%) 25994(34.45%) 300~500 26316(11.75%) 19970(26.47%) 500~1000 28954(12.93%) 13664(18.11%) 1000~2000 46764(20.88%) 8266(10.96%) 2000+ 91650(40.92%) 7558(10.02%) 总数量 223983 75452 总长度 438510672 60036488 N50长度 3096 1543 平均长度 1957.79 795.69 注:括号内数据为各长度区间对应的转录本、unigenes组装比例。 2.2 基因功能注释
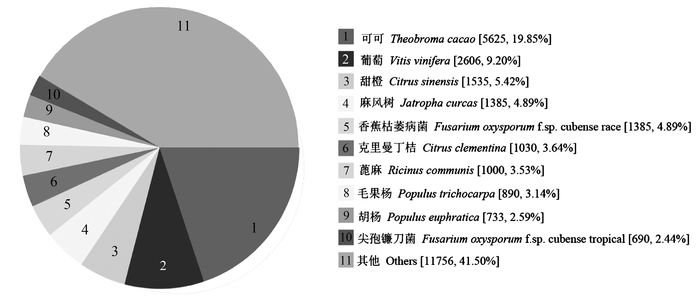
采用Nr、UniProt/Swiss-Prot、GO、COG、KEGG、KOG、Pfam和eggNOG等8个数据库对辣木叶转录组的75 452条unigene进行注释(E值<10-5)(表 2)。结果表明,有40.88%的unigene (30 847条)得到注释,剩余的unigene (59.22%)未能得到注释(表 2)。长度大于1 000 nt的基因中仅有14.24%(2 253条)未得到注释,而小于300 nt的unigene中有72.60%未得到注释。Nr数据库注释结果的E值分布结果表明,得到注释的序列中有54.56%(E值<10-60)的序列与数据库中的序列具有很高的同源性,有33.94%(10-60 <E值<10-15)的序列与数据库中的序列具有较高的同源性。Nr数据库注释结果的物种分布见图 1。辣木叶unigene与可可(19.85%)基因的匹配度最高,其次为葡萄(9.20%)和甜橙(5.42%)。
表 2 辣木叶unigene注释结果Table 2. Summary of functional annotations for the assembled unigenes of Moringa oleifera Lam. leaves数据库 注释基因
数量/条注释基因
比例/%300 nt≤长度
<1000 nt注释
基因数量/条长度≥1000 nt
注释基因
数量/条Nr 28336 37.56 9441 13533 Swiss-Prot 16454 21.81 4690 9638 GO 18277 24.22 5936 9029 COG 9689 12.84 2573 5325 KEGG 10130 13.43 3146 5288 KOG 16850 22.33 5050 8305 Pfam 19541 25.90 5323 11339 eggNOG 28948 38.37 9255 13349 All 30847 40.88 10153 13571 ![]() 图 1 Nr数据库BLAST比对结果的物种分布Figure 1. Species distribution of the BLAST search results in Nr database
图 1 Nr数据库BLAST比对结果的物种分布Figure 1. Species distribution of the BLAST search results in Nr database将辣木叶转录组unigene与GO数据库进行比对,有18 277条unigenes获得至少1条GO注释(表 2)。这些unigenes被分为生物学过程、细胞组分和分子功能3大类和52个亚类(表 3)。细胞组分类中细胞、细胞部分和细胞器3类基因最多,分别为8 160、8 160和6 046条。分子功能类中数量最多的为催化活性和结合分类,分别为9 878和9 483条。生物学过程类中比例最高的为代谢过程和细胞过程,分别为12 587和10 821条。
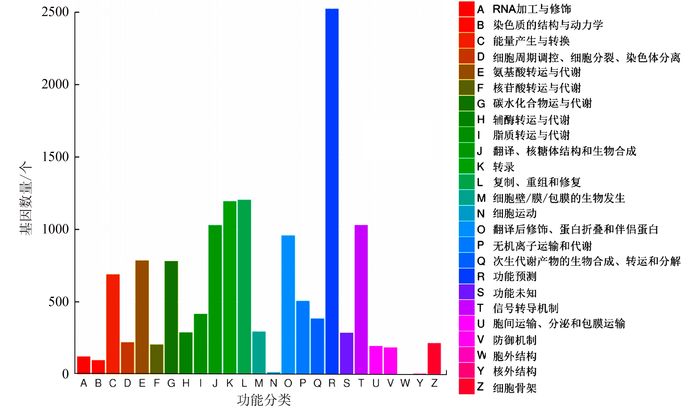
表 3 辣木叶转录组unigene的GO功能分类Table 3. GO classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves生物学过程 细胞组分 分子功能 基因功能 基因数量/个 基因功能 基因数量/个 基因功能 基因数量/个 生殖 265 胞外区 512 蛋白质结合转录因子活性 40 免疫系统过程 254 细胞 8160 核酸结合转录因子活性 500 代谢过程 12587 类核 48 催化活性 9878 细胞过程 10821 膜 4251 受体活性 89 生殖过程 856 病毒体 5 鸟嘌呤核苷酸交换因子活性 46 生物附着 36 细胞连接 293 结构分子活性 650 信号传递 932 胞外基质 7 转运体活性 1296 多细胞组织进程 1495 膜腔 535 结合 9483 发育过程 1602 大分子复合物 2218 电子载体活性 364 生长 306 胞外区 6046 抗氧化活性 145 移动 10 胞外基质组分 1 金属伴侣蛋白活性 4 单组织过程 8955 胞外区域部分 8 酶调节活性 189 生物相 33 细胞器部分 2523 蛋白标签 1 韵律过程 37 病毒体部分 5 翻译调控活性 4 刺激应答 3287 膜部分 2187 营养库活性 20 定位 2855 细胞部分 8160 分子转换器活性 244 多生物过程 759 共质体 292 生物条件 3263 生物组分组织或生物合成 2133 将辣木叶转录组unigene与COG数据库进行比对,共有9 689条unigene得到注释,占总unigene的12.84%(表 2)。COG数据库注释结果如图 2所示,根据功能共分为25类。数量最多的为“功能预测”,共有2 524条,占总数的26.05%。其次为“复制、重组和修复”、“转录”、“翻译、核糖体结构和生物合成”、“信号转导机制”和“翻译后修饰、蛋白折叠和伴侣蛋白”,分别有1 205、1 195、1 030、1 029和958条。“核结构”的数量最少,仅有4条。此外,功能未知的基因有286条,占总数的2.95%。
![]() 图 2 辣木叶转录组unigene的COG功能分类Figure 2. COG classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves
图 2 辣木叶转录组unigene的COG功能分类Figure 2. COG classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves为了更好地了解参与辣木叶生长发育的生物学途径,采用KEGG数据库进行辣木叶转录组unigene注释。10 130条(13.14%) unigenes与KEGG数据库中的序列匹配度高,这些基因分别属于129条KEGG途径(表 4)。其中,3 027条属于代谢途径。代谢途径中碳代谢、氨基酸生物合成和氧化磷酸化途径unigene数量最多,分别为458、399和285条。参与次生代谢物合成的unigene共有407条。
表 4 辣木叶转录组unigene的KEGG功能分类Table 4. KEGG classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves代谢通路 基因数量
/个代谢通路 基因数量
/个代谢通路 基因数量
/个核糖体 517 磷酸肌醇代谢 77 缬氨酸, 亮氨酸和异亮氨酸代谢 34 碳代谢 458 磷脂酰肌醇信号系统 77 叶酸-碳单位循环代谢通路 32 氨基酸生物合成 399 核苷酸切除修复 67 类黄酮生物合成 30 内质网蛋白加工 331 苯丙氨酸,酪氨酸和色氨酸生物合成 64 硒化合物代谢 29 氧化磷酸化 285 β-丙氨酸代谢 63 其他多糖降解 29 剪接体 266 抗坏血酸代谢 62 组氨酸代谢 28 淀粉和蔗糖代谢 255 脂肪酸降解 62 磷酸肌醇生物合成 25 RNA转运 250 萜类骨架生物合成 62 高脚类,二烯类和姜素生物合成 24 糖酵解和糖异生 216 氰基氨基酸代谢 61 花生四烯酸代谢 23 嘌呤代谢 211 ABC转运体 61 硫胺代谢 22 植物激素信号转导 206 RNA聚合酶 61 核黄素代谢 21 内吞作用 197 酪氨酸代谢 60 二萜生物合成 21 mRNA监控通路 195 同源重组 60 柠檬烯、蒎烯降解 20 氨基糖和核苷酸糖代谢 183 脂肪酸生物合成 57 牛磺酸和亚牛磺酸代谢 18 植物病原菌互作 183 丙酸代谢 57 烟酸和烟酰胺代谢 18 吞噬体 176 蛋白质输出 57 生物素代谢 18 泛素介导的蛋白水解 174 色氨酸代谢 55 叶酸生物合成 18 RNA降解 162 DNA复制 55 硫中继系统 18 丙酮酸代谢 160 卟啉和叶绿素代谢 53 维生素B6代谢 16 嘧啶代谢 153 囊泡运输中的陷阱相互作用 52 赖氨酸生物合成 15 苯丙素生物合成 153 不饱和脂肪酸生物合成 51 光合作用-天线蛋白 14 半胱氨酸和蛋氨酸代谢 152 基底转录因子 50 糖胺聚糖降解 14 柠檬酸循环 139 N-多糖生物合成 48 亚油酸代谢 14 光合生物中的碳固定 129 氮代谢 48 玉米素生物合成 14 苯丙氨酸代谢 125 赖氨酸降解 47 酮体合成和降解 13 真核生物核糖体合成 123 α-亚麻酸代谢 47 糖鞘脂类生物合成-globo系列 12 二羧酸代谢 120 碱基切除修复 47 C5分支的二元酸代谢 12 精氨酸和脯氨酸代谢 117 硫代谢 46 芳香化合物降解 11 2-氧代羧酸代谢 116 光合作用 44 倍半萜和三萜类化合物生物合成 10 脂肪酸代谢 115 丁酸代谢 44 芥子油苷生物合成 10 甘油磷脂代谢 107 错配修复 44 油菜素内酯生物合成 9 丙氨酸,阿斯帕特酸和谷氨酸酯代谢 104 脂肪酸延伸 42 非同源性末端接合 9 蛋白酶体 104 自噬调节 41 其他类型o-聚糖生物合成 8 戊糖、葡萄糖醛酸转换 103 类固醇生物合成 40 单萜类生物合成 8 氨酰-tRNA生物合成 103 泛醌和其他萜类生物合成 40 鞘糖脂生物合成 6 过氧物酶体 103 醚脂类代谢 40 咖啡因代谢 5 谷胱甘肽代谢 96 哌啶和吡啶生物碱生物合成 40 花色苷生物合成 4 甘氨酸,丝氨酸和三氨酸代谢 92 植物昼夜节律 40 硫辛酸代谢 3 磷酸戊糖途径 91 泛酸盐和辅酶a生物合成 38 异黄酮生物合成 3 果糖和甘露糖代谢 91 异喹啉生物碱生物合成 37 万古霉素抗药性 3 半乳糖代谢 89 角质,亚氨酸和蜡生物合成 36 黄酮和黄酮醇生物合成 2 缬氨酸, 亮氨酸和异亮氨酸降解 81 鞘脂类代谢 36 碳青霉烯生物合成 1 甘油酯代谢 77 类胡萝卜素生物合成 36 酮化合物糖单位生物合成 1 2.3 基因表达量分析
对unigene的表达量进行分析,发现unigene的FPKM值在0~15 602.35,其中FPKM值大于1 000的unigene有43条,FPKM值介于100~1 000的unigene有779条,FPKM值介于10~100的unigene有8 977条,FPKM值介于1~10的unigene有10 762条。表达量最高的10个基因如表 5所示,其中有7个基因(二磷酸核酮糖羧化酶小链、叶绿体叶绿素a/b结合蛋白、二磷酸核酮糖羧化酶/加氧酶活化酶和铁氧还蛋白)与光合作用有关。根据KEGG注释结果,核糖体相关unigene的FPKM值在0~647.44,其中有214个unigene的FPKM值大于10,FPKM值大于100的unigene有118条,FPKM值最高的为50S核糖体蛋白L12、40核糖体蛋白S4、60S酸性核糖体蛋白P0、50S核糖体蛋白L1和40S核糖体蛋白S14-3。参与氨基酸生物合成途径的unigene中有167条FPKM值大于10,其中FPKM值大于100的unigene有27条,FPKM值最高的依次为果糖-二磷酸醛缩酶、转酮醇酶和磷酸甘油酸激酶基因。参与内质网蛋白加工的unigene中有141条FPKM值大于10。
表 5 辣木叶转录组中表达量最高的10条基因Table 5. The 10 most abundant unigenes in transcriptome of Moringa oleifera Lam. leaves编号 基因名称 FPKM值 c18407.graph_c0 二磷酸核酮糖羧化酶小链 15602.35 c30652.graph_c0 叶绿体叶绿素a/b结合蛋白 12537.82 c35268.graph_c0 二磷酸核酮糖羧化酶小链 8535.69 c33703.graph_c0 类Win蛋白 5152.12 c8930.graph_c0 二磷酸核酮糖羧化酶/加氧酶活化酶 3702.24 c23488.graph_c0 泛素蛋白 2702.34 c30638.graph_c1 氨基环丙烷羧酸氧化酶 2458.66 c35274.graph_c0 叶绿体叶绿素a/b结合蛋白 2268.96 c8931.graph_c0 铁氧还蛋白 2057.79 c35272.graph_c0 叶绿体叶绿素a/b结合蛋白 2011.20 3. 讨论与结论
RNA-seq具有高通量和成本低等诸多优点,已广泛应用于植物功能基因组学研究[26-31]。本研究采用Illumina HiSeq 4000对不同发育时期辣木叶的混合样品进行转录组测序,共组装得到75 452条unigene,为辣木功能基因挖掘提供了大量的基础数据。邓丽婷[19]对成熟辣木叶进行转录组测序仅组装得到45 052条unigene,获得基因信息量明显低于本研究。这可能与本研究采用混合样品进行测序有关。王兴春等[32]也认为采用混合样品进行测序有利于获得更丰富的基因信息。但不排除受测序平台等因素影响。
采用Nr、GO、COG和KEGG等数据库对辣木叶转录组的unigene进行注释,仅有40.88%的unigene获得注释信息。这可能与公共数据库中缺乏辣木及其近缘物种的基因组信息有关。此外,本研究中有72.60%小片段unigene(长度小于300 nt)未得到注释,而长度在1 000 nt以上的unigene注释率达到85.76%。可见,大量短片段序列的存在也是导致注释率较低的重要原因。KEGG代谢途径分析结果表明,辣木叶中有大量核糖体、氨基酸生物合成和内质网蛋白加工等与蛋白质和氨基酸合成相关基因的表达。同时,COG数据库注释结果也表明注释的基因中有大量与蛋白质合成相关的基因(“翻译、核糖体结构和生物合成”和“信号转导机制”和“翻译后修饰、蛋白折叠和伴侣蛋白”)。这些基因的表达为辣木叶高蛋白质和氨基酸含量奠定了基础。除了含有丰富的蛋白质,辣木叶中还含有大量具有生物活性的次生代谢物[5]。次生代谢物的生物合成受大量基因的调控,合成与代谢途径十分复杂。克隆和鉴定次生代谢物合成和调控基因,解析次生代谢途径调控机制,是通过生物工程手段生产次生代谢物的前提。本研究采用公共数据库共注释了30 847条unigene。其中,407条基因参与次生代谢物的合成。这些次生代谢物合成相关基因为解析次生代谢途径及其调控机制提供了丰富的基础数据。
本研究建立了不同发育阶段辣木叶的转录组数据库,组装得到75 452条unigene,并进行功能注释,共有30 847条unigene得到注释。其中,分别有18 277、9 689和129条在GO数据库、COG数据库和KEGG数据库中得到注释。结果表明辣木叶中有大量氨基酸、蛋白质合成和次生代谢产物合成相关的基因表达,这些基因的表达为其高营养价值的形成奠定了物质基础。本研究的结果将为辣木叶营养与药用品质形成相关功能基因挖掘和分子标记开发提供参考,为辣木分子生物学研究与分子育种奠定基础。
-
![]()
图 1 Nr数据库BLAST比对结果的物种分布
Figure 1. Species distribution of the BLAST search results in Nr database
![]()
图 2 辣木叶转录组unigene的COG功能分类
Figure 2. COG classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves
表 1 辣木叶转录组组装结果
Table 1 Summary of sequencing and de novo assembly of Moringa oleifera Lam. leaves
长度区间/nt 转录本/条 unigenes/条 200~300 30299(13.53%) 25994(34.45%) 300~500 26316(11.75%) 19970(26.47%) 500~1000 28954(12.93%) 13664(18.11%) 1000~2000 46764(20.88%) 8266(10.96%) 2000+ 91650(40.92%) 7558(10.02%) 总数量 223983 75452 总长度 438510672 60036488 N50长度 3096 1543 平均长度 1957.79 795.69 注:括号内数据为各长度区间对应的转录本、unigenes组装比例。  下载: 导出CSV
下载: 导出CSV
表 2 辣木叶unigene注释结果
Table 2 Summary of functional annotations for the assembled unigenes of Moringa oleifera Lam. leaves
数据库 注释基因
数量/条注释基因
比例/%300 nt≤长度
<1000 nt注释
基因数量/条长度≥1000 nt
注释基因
数量/条Nr 28336 37.56 9441 13533 Swiss-Prot 16454 21.81 4690 9638 GO 18277 24.22 5936 9029 COG 9689 12.84 2573 5325 KEGG 10130 13.43 3146 5288 KOG 16850 22.33 5050 8305 Pfam 19541 25.90 5323 11339 eggNOG 28948 38.37 9255 13349 All 30847 40.88 10153 13571
下载: 导出CSV
表 3 辣木叶转录组unigene的GO功能分类
Table 3 GO classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves
生物学过程 细胞组分 分子功能 基因功能 基因数量/个 基因功能 基因数量/个 基因功能 基因数量/个 生殖 265 胞外区 512 蛋白质结合转录因子活性 40 免疫系统过程 254 细胞 8160 核酸结合转录因子活性 500 代谢过程 12587 类核 48 催化活性 9878 细胞过程 10821 膜 4251 受体活性 89 生殖过程 856 病毒体 5 鸟嘌呤核苷酸交换因子活性 46 生物附着 36 细胞连接 293 结构分子活性 650 信号传递 932 胞外基质 7 转运体活性 1296 多细胞组织进程 1495 膜腔 535 结合 9483 发育过程 1602 大分子复合物 2218 电子载体活性 364 生长 306 胞外区 6046 抗氧化活性 145 移动 10 胞外基质组分 1 金属伴侣蛋白活性 4 单组织过程 8955 胞外区域部分 8 酶调节活性 189 生物相 33 细胞器部分 2523 蛋白标签 1 韵律过程 37 病毒体部分 5 翻译调控活性 4 刺激应答 3287 膜部分 2187 营养库活性 20 定位 2855 细胞部分 8160 分子转换器活性 244 多生物过程 759 共质体 292 生物条件 3263 生物组分组织或生物合成 2133
下载: 导出CSV
表 4 辣木叶转录组unigene的KEGG功能分类
Table 4 KEGG classifications of unigenes for transcriptome of Moringa oleifera Lam. leaves
代谢通路 基因数量
/个代谢通路 基因数量
/个代谢通路 基因数量
/个核糖体 517 磷酸肌醇代谢 77 缬氨酸, 亮氨酸和异亮氨酸代谢 34 碳代谢 458 磷脂酰肌醇信号系统 77 叶酸-碳单位循环代谢通路 32 氨基酸生物合成 399 核苷酸切除修复 67 类黄酮生物合成 30 内质网蛋白加工 331 苯丙氨酸,酪氨酸和色氨酸生物合成 64 硒化合物代谢 29 氧化磷酸化 285 β-丙氨酸代谢 63 其他多糖降解 29 剪接体 266 抗坏血酸代谢 62 组氨酸代谢 28 淀粉和蔗糖代谢 255 脂肪酸降解 62 磷酸肌醇生物合成 25 RNA转运 250 萜类骨架生物合成 62 高脚类,二烯类和姜素生物合成 24 糖酵解和糖异生 216 氰基氨基酸代谢 61 花生四烯酸代谢 23 嘌呤代谢 211 ABC转运体 61 硫胺代谢 22 植物激素信号转导 206 RNA聚合酶 61 核黄素代谢 21 内吞作用 197 酪氨酸代谢 60 二萜生物合成 21 mRNA监控通路 195 同源重组 60 柠檬烯、蒎烯降解 20 氨基糖和核苷酸糖代谢 183 脂肪酸生物合成 57 牛磺酸和亚牛磺酸代谢 18 植物病原菌互作 183 丙酸代谢 57 烟酸和烟酰胺代谢 18 吞噬体 176 蛋白质输出 57 生物素代谢 18 泛素介导的蛋白水解 174 色氨酸代谢 55 叶酸生物合成 18 RNA降解 162 DNA复制 55 硫中继系统 18 丙酮酸代谢 160 卟啉和叶绿素代谢 53 维生素B6代谢 16 嘧啶代谢 153 囊泡运输中的陷阱相互作用 52 赖氨酸生物合成 15 苯丙素生物合成 153 不饱和脂肪酸生物合成 51 光合作用-天线蛋白 14 半胱氨酸和蛋氨酸代谢 152 基底转录因子 50 糖胺聚糖降解 14 柠檬酸循环 139 N-多糖生物合成 48 亚油酸代谢 14 光合生物中的碳固定 129 氮代谢 48 玉米素生物合成 14 苯丙氨酸代谢 125 赖氨酸降解 47 酮体合成和降解 13 真核生物核糖体合成 123 α-亚麻酸代谢 47 糖鞘脂类生物合成-globo系列 12 二羧酸代谢 120 碱基切除修复 47 C5分支的二元酸代谢 12 精氨酸和脯氨酸代谢 117 硫代谢 46 芳香化合物降解 11 2-氧代羧酸代谢 116 光合作用 44 倍半萜和三萜类化合物生物合成 10 脂肪酸代谢 115 丁酸代谢 44 芥子油苷生物合成 10 甘油磷脂代谢 107 错配修复 44 油菜素内酯生物合成 9 丙氨酸,阿斯帕特酸和谷氨酸酯代谢 104 脂肪酸延伸 42 非同源性末端接合 9 蛋白酶体 104 自噬调节 41 其他类型o-聚糖生物合成 8 戊糖、葡萄糖醛酸转换 103 类固醇生物合成 40 单萜类生物合成 8 氨酰-tRNA生物合成 103 泛醌和其他萜类生物合成 40 鞘糖脂生物合成 6 过氧物酶体 103 醚脂类代谢 40 咖啡因代谢 5 谷胱甘肽代谢 96 哌啶和吡啶生物碱生物合成 40 花色苷生物合成 4 甘氨酸,丝氨酸和三氨酸代谢 92 植物昼夜节律 40 硫辛酸代谢 3 磷酸戊糖途径 91 泛酸盐和辅酶a生物合成 38 异黄酮生物合成 3 果糖和甘露糖代谢 91 异喹啉生物碱生物合成 37 万古霉素抗药性 3 半乳糖代谢 89 角质,亚氨酸和蜡生物合成 36 黄酮和黄酮醇生物合成 2 缬氨酸, 亮氨酸和异亮氨酸降解 81 鞘脂类代谢 36 碳青霉烯生物合成 1 甘油酯代谢 77 类胡萝卜素生物合成 36 酮化合物糖单位生物合成 1
下载: 导出CSV
表 5 辣木叶转录组中表达量最高的10条基因
Table 5 The 10 most abundant unigenes in transcriptome of Moringa oleifera Lam. leaves
编号 基因名称 FPKM值 c18407.graph_c0 二磷酸核酮糖羧化酶小链 15602.35 c30652.graph_c0 叶绿体叶绿素a/b结合蛋白 12537.82 c35268.graph_c0 二磷酸核酮糖羧化酶小链 8535.69 c33703.graph_c0 类Win蛋白 5152.12 c8930.graph_c0 二磷酸核酮糖羧化酶/加氧酶活化酶 3702.24 c23488.graph_c0 泛素蛋白 2702.34 c30638.graph_c1 氨基环丙烷羧酸氧化酶 2458.66 c35274.graph_c0 叶绿体叶绿素a/b结合蛋白 2268.96 c8931.graph_c0 铁氧还蛋白 2057.79 c35272.graph_c0 叶绿体叶绿素a/b结合蛋白 2011.20
下载: 导出CSV
-
[1] 党选民, 曹振木, 杨(龙天), 等.热带珍稀植物辣木的特性及其开发利用[J].中国农学通报, 2004, 20(20):133-136. http://cpfd.cnki.com.cn/Article/CPFDTOTAL-ZGNX200400001033.htm [2] ANWAR F, LATIF S, ASHRAF M, et al.Moringa oleifera:a food plant with multiple medicinal uses[J]. Phytotherapy Research, 2007, 21(1):17-25. DOI: 10.1002/(ISSN)1099-1573
[3] MISHRA S, SINGH P, SINGH S. Nutritional and medicinal value of Moringa oleifera leaves:potential and prospects[J]. Forestry Bulletin, 2011, 11(1):46-58. http://downloads.hindawi.com/journals/jt/2014/786979.xml
[4] SUJATHA B K, POONAM P. Moringa Oleifera-Nature's Gold[J]. Imperial Journal of Interdisciplinary Research, 2017, 3(5):1175-1179. DOI: 10.1556/1646.8.2016.3.7
[5] MOYO B, MASIKA P, HUGO A, et al. Nutritional characterization of Moringa (Moringa oleifera Lam.) leaves[J]. African Journal of Biotechnology, 2011, 10(60):12925-12933. DOI: 10.5897/AJB
[6] 刘凤霞, 王苗苗, 赵有为, 等.辣木中功能性成分提取及产品开发的研究进展[J].食品科学, 2015, 36(19):282-286. DOI: 10.7506/spkx1002-6630-201519051 [7] MISHRA S P, SINGH P, SINGH S. Processing of Moringa oleifera leaves for human consumption[J]. Bulletin of Environment, Pharmacology and Life Sciences, 2012, 2(1):28-31. http://downloads.hindawi.com/journals/jt/2014/786979.xml
[8] FAHEY J W. Moringa oleifera:a review of the medical evidence for its nutritional, therapeutic, and prophylactic properties. part 1[J]. Trees for life Journal, 2005, (1):5. DOI: 10.1128/jmbe.v18i3.1306
[9] 彭磊, 田洋, 解静, 等.世界辣木发展现状及市场前景分析[J].世界农业, 2015, (9):143-146. http://d.wanfangdata.com.cn/Periodical_shijny201505027.aspx [10] WATERMAN C, CHENG D M, ROJAS-SILVA P, et al. Stable, water extractable isothiocyanates from Moringa oleifera leaves attenuate inflammation in vitro[J]. Phytochemistry, 2014, 103(Supplement C):114-122. https://es.scribd.com/document/119809815/JMPR-15-August-2012-Issue
[11] JUNG I L. Soluble extract from Moringa oleifera leaves with a new anticancer activity[J]. PLoS ONE, 2014, 9(4):e95492. DOI: 10.1371/journal.pone.0095492
[12] RICHTER N, SIDDHURAJU P, BECKER K. Evaluation of nutritional quality of moringa (Moringa oleifera Lam.) leaves as an alternative protein source for Nile tilapia (Oreochromis niloticus L.)[J]. Aquaculture, 2003, 217(1):599-611. http://www.ingentaconnect.com/content/els/00448486/2003/00000217/00000001/art00497
[13] 刘子记, 孙继华, 刘昭华, 等.特色植物辣木的应用价值及发展前景分析[J].热带作物学报, 2014, 35(9):1871-1878. http://www.cnki.com.cn/Article/CJFDTotal-RDNK201506003.htm [14] 吴崸, 蔡志华, 魏烨昕, 等.辣木作为新型植物性蛋白质饲料的研究进展[J].动物营养学报, 2013, 25(3):503-511. http://www.docin.com/p-747955573.html [15] TIAN Y, ZENG Y, ZHANG J, et al. High quality reference genome of drumstick tree (Moringa oleifera Lam.), a potential perennial crop[J]. Science China Life Sciences, 2015, 58(7):627-638. DOI: 10.1007/s11427-015-4872-x
[16] 田洋, 董扬, 盛军, 等.辣木转录因子家族初步鉴定与分析[J].云南农业大学学报:自然科学版, 2016, 31(6):450-461. http://www.cqvip.com/QK/94499X/201603/668956714.html [17] KONDO T, FUJIKAWA Y, UEDA A, et al. Cloning and gene expression analysis of ascorbic acid biosynthesis enzymes in Moringa oleifera[J]. African Journal of Agricultural Research, 2015, 10(22):2274-2285. DOI: 10.5897/AJAR
[18] DENG L T, WU Y L, LI J C, et al. Screening reliable reference genes for RT-qPCR analysis of gene expression in Moringa oleifera[J]. PLoS ONE, 2016, 11(8):e0159458. DOI: 10.1371/journal.pone.0159458
[19] 邓丽婷. 基于转录组测序的辣木内参基因筛选和三个代谢途径相关基因的表达分析[D]. 广州: 华南农业大学, 2016: 48-49. http://cdmd.cnki.com.cn/Article/CDMD-10564-1016923469.htm [20] FANG Z Z, ZHOU D R, YE X F, et al. Identification of candidate anthocyanin-related genes by transcriptomic analysis of 'Furongli' plum (Prunus salicina Lindl.) during fruit ripening using RNA-Seq[J]. Frontiers in Plant Science, 2016, 7(166):1338. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5005409/
[21] GRABHERR M G, HAAS B J, YASSOUR M, et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome[J]. Nature Biotechnology, 2011, 29(7):644-652. DOI: 10.1038/nbt.1883
[22] LANGMEAD B, TRAPNELL C, POP M, et al. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome[J]. Genome Biology, 2009, 10(3):1-10. http://www.oalib.com/paper/185058
[23] LI B, DEWEY C. RSEM:accurate transcript quantification from RNA-Seq data with or without a reference genome[J]. BMC Bioinformatics, 2011, 12(1):323. DOI: 10.1186/1471-2105-12-323
[24] ONESA A, GÖTZ S, GARCÍA-GÓMEZ J M, et al. Blast2GO:a universal tool for annotation, visualization and analysis in functional genomics research[J]. Bioinformatics, 2005, 21(18):3674-3676. DOI: 10.1093/bioinformatics/bti610
[25] YE J, FANG L, ZHENG H, et al. WEGO:a web tool for plotting GO annotations[J]. Nucleic Acids Res, 2006, 34(S2):W293-W297. https://academic.oup.com/nar/article/34/suppl_2/W293/2505468/WEGO-a-web-tool-for-plotting-GO-annotations
[26] 张少平, 邱珊莲, 郑云云, 等.紫色黄秋葵转录组功能基因测序及分析[J].核农学报, 2017, 31(4):643-653. DOI: 10.11869/j.issn.100-8551.2017.04.0643 [27] 向君亮, 黄殷.薏苡幼苗叶片转录组分析[J].农业生物技术学报, 2017, 25(3):386-396. http://www.jabiotech.org/CN/abstract/abstract10663.shtml [28] 武红霞, 许文天, 罗纯, 等.芒果果实转录组数据组装及基因功能注释简[J].热带作物学报, 2016, 37(11):2191-2198. DOI: 10.3969/j.issn.1000-2561.2016.11.025 [29] 蔡年辉, 邓丽丽, 许玉兰, 等.基于高通量测序的云南松转录组分析[J].植物研究, 2016, 36(1):75-83. DOI: 10.7525/j.issn.1673-5102.2016.01.011 [30] 姜福星, 杨丽娟, 高素萍, 等.碰碰香叶片的转录组测序分析[J].分子植物育种, 2015, 13(10):2310-2319. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=fzzw201510025&dbname=CJFD&dbcode=CJFQ [31] 吴超, 彭娟, 向林, 等.基于高通量测序的铁皮石斛叶片转录组分析[J].分子植物育种, 2012, 14(12):3334-3346. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=fzzw201612009&dbname=CJFD&dbcode=CJFQ [32] 王兴春, 谭河林, 陈钊, 等.基于RNA-Seq技术的连翘转录组组装与分析及SSR分子标记的开发[J].中国科学:生命科学, 2015, 45(3):301-310. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=jcxk201503008&dbname=CJFD&dbcode=CJFQ -
期刊类型引用(0)
其他类型引用(1)
计量
- 文章访问数: 2032
- HTML全文浏览量: 243
- PDF下载量: 130
- 被引次数: 1
