

Vector Construction and Immunogenicity of S and N Gene DNA Vaccine for TGEV
-
摘要:目的
构建猪传染性胃肠炎病毒(transmissible gastroenteritis virus, TGEV)S、N基因的DNA疫苗载体,并进行免疫原性试验,为猪传染性胃肠炎(transmissible gastroenteritis, TGE)的防控和DNA疫苗研究提供技术支撑和基础数据。
方法扩增S基因的A位点、D位点和N基因,并将N基因(单独)、A位点和D位点(融合)克隆至pCDNA3.1-His-C构建重组疫苗载体,运用生物信息学软件预测分析S(A-D)蛋白、N蛋白二级结构组成、三级构像、亚细胞定位和优势B细胞抗原表位。将构建成功的重组载体分别转染至PK-15细胞进行间接免疫荧光试验,运用共聚焦检测重组蛋白的表达分布情况。将重组疫苗载体单独或联合免疫小鼠,运用间接ELISA检测IgG抗体水平。
结果扩增出S基因的A位点、D位点和N基因,大小分别为498、606、
1149 bp。构建了A位点与D位点(融合)、N基因(单独)的DNA疫苗重组载体p-S(A-D)-His和p-N-His。生物信息学软件预测分析发现TGEV感染宿主细胞时N蛋白主要定位于细胞核和线粒体,S(A-D)蛋白主要定位于细胞质和线粒体,S(A-D)蛋白具有7个优势B细胞抗原表位,N蛋白具有8个优势B细胞抗原表位。重组载体p-S(A-D)-His和p-N-His均在PK-15细胞内成功表达,且S(A-D)-His和N-His在PK-15细胞核和细胞质中均有分布。重组疫苗载体免疫小鼠后,免疫效果由高至低依次为p-N-His>p-S(A-D)-His + p-N-His>p-S(A-D)-His。结论本研究构建了TGEV的 S、N基因的DNA疫苗载体,免疫小鼠后均产生了较强的特异性抗体,为TGEV的核酸疫苗的研制提供了基础材料和依据。
Abstract:ObjectiveDNA vaccine vector of S and N genes of porcine transmissible gastroenteritis virus (TGEV) was constructed with the vaccine immunogenicity determined to pave the way for studying, preventing, and controling TGE.
MethodA and D sites on S and N from a TGEV were amplified. The N gene alone as well as the A and D sites fusion were cloned into the vaccine vector pCDNA3.1-His-C. Bioinformatics software was used to predict and analyze the secondary structure, tertiary configuration, subcellular localization, and dominant B cell epitope of S (A-D) and N proteins. The recombinant vectors were transfected into PK-15 cells, and expression distribution of N and the A and D sites fusion detected by indirect immunofluorescence and confocal detection. Mice were immunized with the single or combined recombinant vaccine vector to detect the IgG antibody using indirect ELISA.
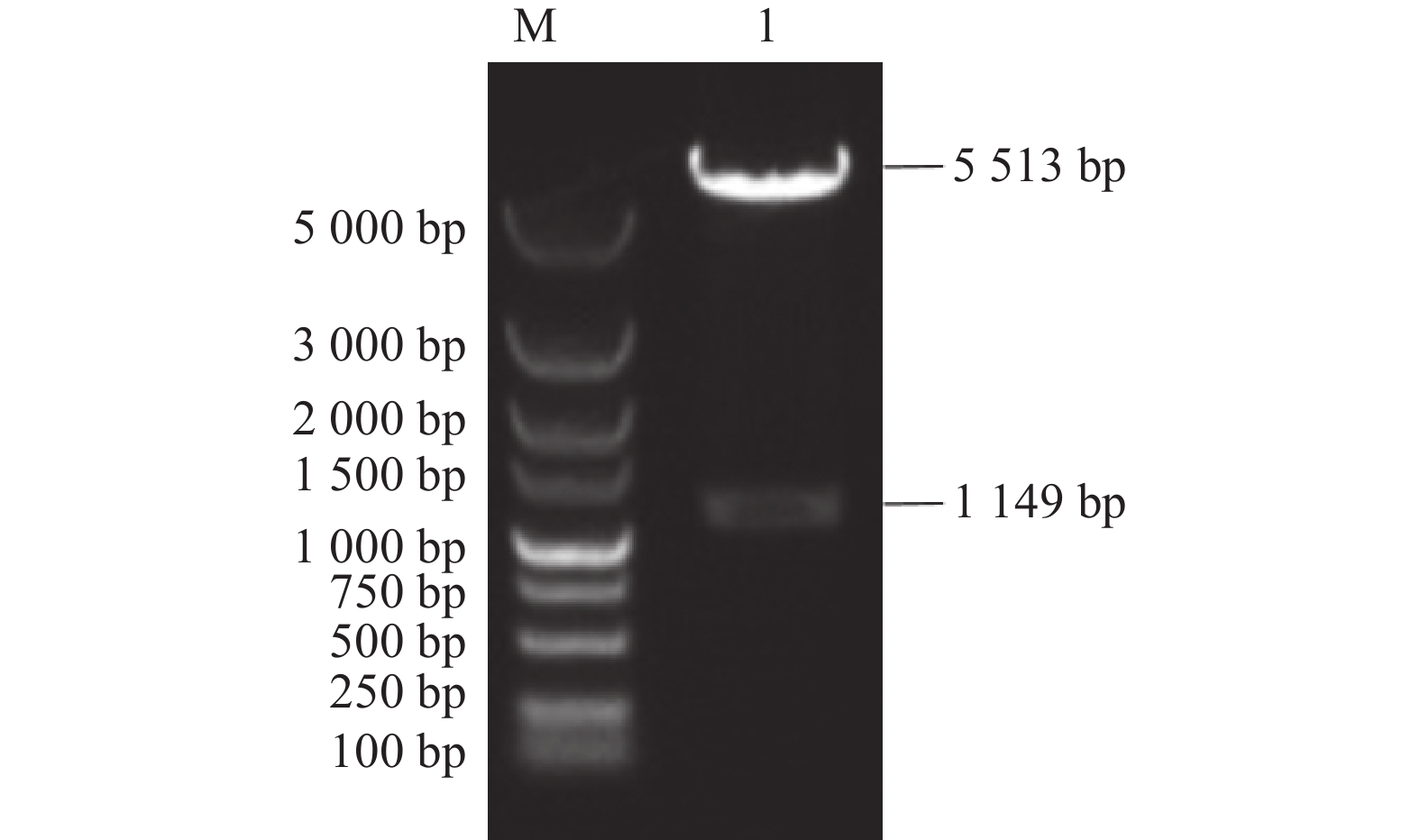
ResultThe A and D sites of the S were 498 bp and 606 bp, respectively, and the N,
1149 bp in length. The nucleic acid vaccine expression vectors p-S (A-D)-His and p-N-His for the A and D sites (fusion) and N were constructed. Bioinformatics software predicted that, when TGEV infected the host cells, N protein was mainly located in the nucleus and mitochondria and S (A-D) largely in the cytoplasm and mitochondria, while S (A-D) had 7 and N, 8 dominant B cell epitopes. All p-S (A-D)-His and p-N-His were successfully expressed in PK-15 cells distributed in the nucleus and cytoplasm. The immunized mice showed an effect of immunity in the order of p-N-His>p-S (A-D)-His + p-N-His>p-S (A-D)-His.ConclusionThe DNA vaccine vectors of S and N of TGEV were successfully constructed. Strong specific antibodies were generated in lab mice after the immunization.
-
0. 引言
【研究意义】植物种质资源的收集与保存是植物研究重要内容,其中超低温保存作为植物组织和细胞长期保存的理想方法,尤其适合营养繁殖作物的茎尖或分生组织的保存,具有直接再生完整小植株、减少遗传变异等优点[1]。海棠是蔷薇科(Rosaceae)苹果属(Malus)中果径较小(≤5 cm)的落叶乔木或小乔木,其中观赏海棠(Malus sp.)观赏价值高,是重要的优质绿化树种,研究海棠茎尖超低温保存在生产上具有重要意义。【前人研究进展】超低温保存后较高的存活率和再生率是主要的技术目标,已有研究显示,玻璃化超低温保存中活性氧(Reactive oxygen species, ROS)诱导的氧化应激是引起植物材料冻存后存活率下降的原因之一,因此,可以采用外源抗氧化剂来保护机体在超低温保存中免受伤害,如过氧化氢酶(Catalase, CAT)[2]、超氧化物歧化酶(Superoxide dismutase, SOD)[3]等酶类抗氧化剂以及抗坏血酸(Ascorbic acid, AsA)[4]和谷胱甘肽(Glutathione, GSH)[5]等非酶类抗氧化剂,均可以有效降低ROS生成量,减少氧化应激的发生,显著改善超低温保存效果。此外,细胞程序性死亡(Programmed cell death, PCD)作为一种基因编码主动性的死亡方式,在植物超低温保存中也扮演了重要的角色,添加细胞凋亡抑制剂D-CHO[6]、NO供体(Sodium Nitroprusside, SNP)[7]和乙烯(乙烯利Ethephon, Eth)[8] 等抑制PCD发生的物质可以提高存活率。【本研究切入点】目前除了对苹果属一些种类有少量超低温保存[9-12],特别是涉及观赏海棠的超低温保存研究极少,而极具观赏价值的北美海棠品种之一——红丽海棠(Malus Red Splendor)超低温保存鲜见报道。【拟解决的关键问题】本文以红丽海棠茎尖为试材,研究其超低温保存技术程序,探讨添加不同含量的抗氧化剂及PCD抑制剂对茎尖超低温保存冻后存活率的影响,建立红丽海棠玻璃化超低温保存技术程序,为观赏海棠种质资源保存提供一种技术思路,并为抗氧化剂和PCD抑制剂在超低温保存中的应用提供参考。
1. 材料与方法
1.1 试验材料
供试材料为观赏海棠品种红丽,以健康且带有饱满腋芽的当年生枝条为外植体(采自国家植物园海棠栒子园),在1/2MS+1.5 mg·L−1 6BA+0.1 mg·L−1 IAA的培养基中诱导无菌苗,并于1/2MS+1.0 mg·L−1 6-BA+0.5 mg·L−1 IAA的培养基中进行继代培养,诱导和继代培养条件:(23±3)℃,光照强度40 μmol·m−2·s−1,光照时间14 h·d−1,取生长旺盛的组培苗用于超低温保存试验。
1.2 试验方法
1.2.1 茎尖玻璃化法超低温保存
根据初步预试验设置玻璃化超低温保存的基本程序,再采用“逐步单因子法”对关键环节进行优化。(1)预培养:将红丽组培苗切成5 mm左右的带顶芽茎段,分别接种在不同蔗糖浓度(0.1、0.3、0.5、0.7、0.9 mol·L−1)的MS培养基中4 ℃下培养2 d,然后继续下述程序:(2)装载(Loading):在双目解剖镜下,用镊子和手术刀切下带3~4片叶原基、大小为1.5~2.0 mm的茎尖;将切下的茎尖放入1.5 mL 装有1 mL MS溶液的离心管中,吸出离心管中的MS溶液,加入1 mL Loading溶液(2 mol·L−1丙三醇+0.4 mol·L−1蔗糖,蒸馏水配置,pH 5.8),在室温下静置处理30 min;(3)PVS2处理:吸出离心管中的Loading溶液,加入1mL PVS2溶液[30%(m/V)丙三醇+15 %(m/V)乙二醇+15%(m/V)二甲基亚砜+0.4 mol·L−1蔗糖,1/2MS盐溶液配置,pH 5.8],在0 ℃于冰浴中处理90 min;(4)液氮冻存:更换新的1 mL PVS2溶液后,封紧离心管口,立即投入液氮,冻存1 h;(5)化冻:将装有PVS2和茎尖的离心管从液氮中取出,迅速放入38 ℃水浴中化冻1 min;(6)去装载(Unloading):将离心管中的PVS2溶液吸出,加入Unloading溶液(用1/2MS盐溶液配置的1.2 mol·L−1蔗糖,pH 5.8)后在室温下震荡洗涤2次,每次10 min。然后用于存活率或恢复生长率测定。相关步骤示意图见图1。
![]() 图 1 茎尖玻璃化超低温保存及存活率检测A.红丽海棠组培苗;B.预培养;C.切取1.5~2.5 mm茎尖;D.茎尖在离心管中;E.经TTC染色变红(左)与未染色(右)的茎尖;F.冻后茎尖接种到恢复培养基中。Figure 1. Survival rate of post-cryopreservation shoot tipsA: Crabapple tissue culture; B: pre-culture; C: 1.5-2.5 mm cut of shoot tip; D: shoot tip segment in centrifugal tube; E: TTC stained red (left) and original (right) shoot tips; F: post-frozen shoot tips inoculated on recovery medium.
图 1 茎尖玻璃化超低温保存及存活率检测A.红丽海棠组培苗;B.预培养;C.切取1.5~2.5 mm茎尖;D.茎尖在离心管中;E.经TTC染色变红(左)与未染色(右)的茎尖;F.冻后茎尖接种到恢复培养基中。Figure 1. Survival rate of post-cryopreservation shoot tipsA: Crabapple tissue culture; B: pre-culture; C: 1.5-2.5 mm cut of shoot tip; D: shoot tip segment in centrifugal tube; E: TTC stained red (left) and original (right) shoot tips; F: post-frozen shoot tips inoculated on recovery medium.关键环节的优化:在获得适宜预培养蔗糖浓度基础上,分别设置不同预处理时间(1、2、3 d),不同Loading溶液处理时间(0、10、20、30、40 min)和不同PVS2溶液处理时间(0、30、60、90、120 min),每个优化环节之前的步骤采用上一步已优化的处理方法,之后的步骤采用上述基本程序,直至完成所有关键环节的优化。每处理10~15个茎尖,重复3次。
1.2.2 存活率测定
采用氯化三苯基四氮唾(TTC)法检测冻存后茎尖的存活率。将经过Unloading溶液处理的茎尖置于含0.4%(m/V)的TTC溶液的离心管中,再放入37 ℃的水浴中黑暗处理30 min,将茎尖取出后,置于体视显微镜下,观察其染色情况。顶端分生组织被染红的茎尖记为存活的茎尖,而未被染红的茎尖记为死亡的茎尖。存活率/%=(被染红的茎尖/所有经过TTC处理的茎尖)×100%。每处理10~15个茎尖,重复3次。
1.2.3 恢复生长率测定
将经过液氮保存后的茎尖接种于MS培养基中暗培养2周后,然后转入正常光照条件(与组织培养的条件相同)下培养,茎尖长叶记为恢复生长。每处理10~15个茎尖,重复3次。
1.2.4 外源抗氧化剂和PCD抑制剂的添加
在建立的玻璃化超低温保存程序的基础上,根据课题组前人研究结果[4, 7-8, 13],分别在预培养基中添加Eth(100、200、400、800 mg·L−1);在Loading溶液中添加CAT,使其终浓度分别为50、100、200、400 U·mL−1和硝普钠(SNP),使其终浓度分别为50、100、200、1000 μmol·L−1;在PVS2溶液中添加GSH,使其终浓度分别为0.04、0.08、0.16、0.32 μmol·L−1;在Unloading溶液中添加AsA,使其终浓度分别为100、200、400、600 μmol·L−1,各处理均以原溶液为对照。
1.2.5 数据处理及分析
采用SPSS 26.0和Excel软件分别对试验数据进行单因素方差分析和数据整理、图表绘制。
2. 结果与分析
2.1 红丽海棠茎尖玻璃化超低温保存程序的建立
2.1.1 预培养基蔗糖浓度对液氮冻存后茎尖存活率的影响
将红丽海棠带顶芽茎段分别在含0.1、0.3、0.5、0.7、0.9 mol·L−1蔗糖的预培养基上进行2 d的培养,之后采用后续的基本程序,测定液氮冻存后茎尖存活率。如图2所示,茎尖冻后存活率随蔗糖浓度的升高先上升再下降,0.3 、0.5 、0.7 mol·L−1蔗糖浓度培养的茎尖存活率没有显著差异,但0.7 mol·L−1蔗糖浓度的预培养基上茎尖冻后存活率最高,为76.54%,显著高于0.1 、0.9 mol·L−1蔗糖浓度。因此红丽海棠超低温保存程序中预培养基的最适蔗糖浓度为0.7 mol·L−1,浓度过低过高则会降低茎尖的存活率。
![]() 图 2 预培养基蔗糖浓度对液氮冻存后茎尖存活率的影响不同小写字母表示不同差异显著(P<0.05),下同。Figure 2. Effect of sucrose concentration in pretreatment medium on survival rate of post-cryopreservation shoot tipsDifferent lowercase letters indicate different significant differences (P<0.05), same as below.
图 2 预培养基蔗糖浓度对液氮冻存后茎尖存活率的影响不同小写字母表示不同差异显著(P<0.05),下同。Figure 2. Effect of sucrose concentration in pretreatment medium on survival rate of post-cryopreservation shoot tipsDifferent lowercase letters indicate different significant differences (P<0.05), same as below.2.1.2 预处理时间对液氮冻存后茎尖存活率的影响
将带顶芽的红丽海棠茎段接种于含0.7 mol·L−1蔗糖的预培养基上,在4 ℃条件下分别培养1 、2 和3 d,继续超低温保存基本程序的后续环节,液氮冻存后红丽茎尖存活率见图3。随预处理时间的增加,茎尖存活率逐渐升高,在2 d时存活率最高达到79.24%,后又下降。表明液氮冻后茎尖存活率受预培养环节冷锻炼时间的影响,适宜时长可以提高茎尖的抗冻能力。
![]() 图 3 预处理时间对液氮冻存后茎尖存活率的影响Figure 3. Effect of pretreatment time on survival rate of post-cryopreservation shoot tips
图 3 预处理时间对液氮冻存后茎尖存活率的影响Figure 3. Effect of pretreatment time on survival rate of post-cryopreservation shoot tips2.1.3 Loading溶液处理时间对液氮冻存后茎尖存活率的影响
红丽海棠茎段在含0.7 mol·L−1蔗糖浓度的预培养2 d后,将茎尖分别在Loading溶液中处理不同时间,继续超低温保存基本程序的后续环节,从液氮取出化冻后茎尖存活率的结果如图4所示。Loading处理20 min时茎尖存活率最高,为60.61%,而随着处理时间的继续延长逐渐下降,处理40 min时的茎尖存活率为0。因此,Loading溶液处理时间不宜过长,否则可能会造成茎尖过度脱水而降低存活率。红丽海棠茎尖超低温保存的适宜Loading处理时间为20 min。
![]() 图 4 Loading溶液处理时间对液氮冻存后茎尖存活率的影响Figure 4. Effect of loading solution treatment time on survival rate of post-cryopreservation shoot tips
图 4 Loading溶液处理时间对液氮冻存后茎尖存活率的影响Figure 4. Effect of loading solution treatment time on survival rate of post-cryopreservation shoot tips2.1.4 PVS2溶液处理时间对液氮冻存后茎尖存活率的影响
红丽海棠茎段在4 ℃下,含0.7 mol·L−1蔗糖浓度的预培养基上培养2 d后,Loading溶液处理20 min,分别采用PVS2溶液处理不同时间,继续超低温保存基本程序的后续环节,结果如图5。未经PVS2溶液处理的茎尖全部死亡,说明PVS2处理对玻璃化超低温保存过程起着至关重要的作用,随处理时间的增加,茎尖冻后存活率逐渐升高,处理90 min时茎尖存活率最高,为62.88%,后随时间的延长而显著下降。红丽海棠茎尖超低温保存PVS2处理适宜时间为90 min。
![]() 图 5 PVS2溶液处理时间对液氮冻存后茎尖存活率的影响Figure 5. Effect of PVS2 treatment time on survival rate of post-cryopreservation shoot tips
图 5 PVS2溶液处理时间对液氮冻存后茎尖存活率的影响Figure 5. Effect of PVS2 treatment time on survival rate of post-cryopreservation shoot tips2.2 外源添加剂对茎尖超低温保存效果的影响
2.2.1 预培养环节添加外源Eth对液氮冻存后茎尖存活率的影响
在红丽海棠茎尖超低温保存中预培养环节添加4个质量浓度的外源Eth,继续超低温保存优化后的后续环节,茎尖冻后存活率见图6。添加100~400 mg·L−1与对照没有显著差异,200 mg·L−1时存活率稍高,而100 、400 mg·L−1时存活率稍低,而质量浓度过高时(800 mg·L−1)存活率显著下降。说明在预培养环节导入200 mg·L−1Eth时可能起到一定保护作用,小幅度提高冻后存活率,高浓度反而会导致其存活率降低。
![]() 图 6 外源Eth对液氮冻存后茎尖存活率的影响Figure 6. Effect of added Eth on survival rate of post-cryopreservation shoot tips
图 6 外源Eth对液氮冻存后茎尖存活率的影响Figure 6. Effect of added Eth on survival rate of post-cryopreservation shoot tips2.2.2 Loading环节添加外源CAT或SNP对液氮冻存后茎尖存活率的影响
采用预培养优化方案后,在Loading环节添加不同浓度的外源CAT,之后继续超低温保存优化后的后续环节,茎尖液氮冻存后存活率见图7。添加200 U·mL−1时,茎尖的存活率最高,为86.67%,显著高于其他处理;含量为50 U·mL−1稍有提高,但差异不显著,其余含量显著下降。由此可知,Loading环节添加适宜浓度的外源CAT浓度可以发挥抗氧化保护作用,显著提高茎尖冻后存活率。
![]() 图 7 外源CAT对液氮冻存后茎尖存活率的影响Figure 7. Effect of added CAT on survival rate of post-cryopreservation shoot tips
图 7 外源CAT对液氮冻存后茎尖存活率的影响Figure 7. Effect of added CAT on survival rate of post-cryopreservation shoot tips采用预培养优化方案后,在Loading环节添加不同浓度的外源SNP,之后继续超低温保存优化后的后续环节,茎尖超低温保存后存活率如图8所示。各浓度处理后冻后存活率均下降,但添加100和1000 μmol·L−1SNP时与对照无明显差异,而添加50和200 μmol·L−1SNP时存活率显著下降,相比对照分别降低了23.25%和13.25%。表明对PCD有拮抗作用的SNP在这里没有显示其作用。
![]() 图 8 外源SNP对液氮冻存后茎尖存活率的影响Figure 8. Effect of added SNP on survival rate of post-cryopreservation shoot tips
图 8 外源SNP对液氮冻存后茎尖存活率的影响Figure 8. Effect of added SNP on survival rate of post-cryopreservation shoot tips2.2.3 PVS2环节添加外源GSH对液氮冻存后茎尖存活率的影响
采用优化的预培养和Loading方案后,在PVS2环节添加不同浓度的外源GSH,之后继续超低温保存后优化的后续环节,茎尖液氮冻存后存活率见图9。添加各种浓度的GSH,较对照均显著提高了茎尖冻后的存活率,其中导入浓度为0.04 μmol·L−1时存活率最高,为94.19%。由此表明,在PVS2环节导入外源GSH可有效发挥抗氧化保护作用,提高存活率。
![]() 图 9 外源GSH对液氮冻存后茎尖存活率的影响Figure 9. Effect of added GSH on survival rate of post-cryopreservation shoot tips
图 9 外源GSH对液氮冻存后茎尖存活率的影响Figure 9. Effect of added GSH on survival rate of post-cryopreservation shoot tips2.2.4 Unloading环节添加外源AsA对液氮冻存后茎尖存活率的影响
采用优化的保存程序后,在Unloading环节导入不同浓度的外源AsA,茎尖超低温保存后存活率如图10所示,除200 μmol·L−1存活率下降外,其余浓度均可提高冻后存活率,效果最好的浓度为400 μmol·L−1,说明外源AsA起到了一定的抗氧化保护作用,小幅度提高了存活率。
![]() 图 10 外源AsA对液氮冻存后茎尖存活率的影响Figure 10. Effect of added AsA on survival rate of post-cryopreservation shoot tips
图 10 外源AsA对液氮冻存后茎尖存活率的影响Figure 10. Effect of added AsA on survival rate of post-cryopreservation shoot tips2.3 红丽海棠茎尖超低温保存后恢复培养
采用优化后的超低温保存程序,在PVS2环节添加0.04 μmol·L−1的外源GSH,之后继续超低温保存后优化的后续环节,并进行恢复培养。没有添加GSH的对照组恢复生长率为16.67%,添加0.04 μmol·L−1外源GSH的恢复生长率为41.39%,提高了24.73%。
3. 讨论
冷锻炼能够提高植物茎尖的抗冻能力[14],通过在含有一定浓度的高糖、山梨醇或DMSO的培养基中培养植物茎尖,可以显著提高冻存后茎尖的存活率[15]。在大部分苹果属玻璃化超低温保存中,都至少需要3~4周的4 ℃或5 ℃的冷锻炼[16-19]。本试验通过4 ℃预培养,起到冷锻炼作用,预培养2 d,超低温保存后的茎尖存活率可到79.24%。在苹果属其他茎尖超低温保存的研究中,有不使用Loading处理直接脱水处理[20-22],本试验不经Loading溶液处理也可达到36.11%的存活率,但经Loading溶液处理20 min的茎尖冻后存活率更高,因为装载阶段是实现玻璃化而脱水处理的过渡阶段,能够避免高浓度玻璃化溶液引起的剧烈渗透变化导致的材料损伤。另外在本试验中0 ℃条件下PVS2处理90 min后茎尖冻后存活率最高,也有研究在室温下进行脱水处理,如将苹果品种嘎啦、望山红在室温下用PVS2处理30 min[10]或40 min[11]后再生率分别为55.6%和17.9%,但Uragami等[23]认为低温能降低玻璃化溶液的高渗作用导致的植物损伤,因此在大多数植物的超低温研究中,玻璃化步骤均在0 ℃下进行[24, 25]。
ROS是需氧生物中细胞有氧代谢的副产物,在正常状态下,机体可以在ROS的产生和清除之间保持动态平衡,但当ROS在逆境胁迫下产生过量时,自身的抗氧化系统,包括CAT、SOD和POD等酶类,以及AsA和GSH等非酶类系统会协同作用清除抵抗ROS可能带来的伤害,但当抗氧化能力不足以清除过量的ROS时,机体就会出现氧化应激。这启发研究人员们通过导入外源抗氧化剂来抵御氧化损伤,研究证明外源CAT[4, 13, 26]、AsA[4, 27]和GSH[6]均有抑制氧化应激的作用。本研究结果也显示,外源抗氧化剂CAT、AsA和GSH可以提高红丽海棠茎尖冻后存活率,说明外源抗氧化剂可能抑制了ROS造成的氧化损伤。3种外源抗氧化剂在不同环节中使用,分别提高了红丽海棠茎尖冻后存活率20.28%、6.75%、27.61%。
PCD作为一种自发性、有序的细胞死亡方式,受基因调控,是生物生活的基本过程。多项研究表明植物PCD存在于植物的各个生长发育过程及植物对非生物胁迫的响应中[28]。有研究显示超低温保存中预培养是PCD的一个信号起始环节,PCD相关调控基因显著上调[7],由于细胞凋亡存在一定的延迟性,预培养后的Loading环节也成为超低温保存中PCD研究的另一重要环节。本试验在预培养和Loading环节添加了PCD抑制剂,结果显示预培养环节添加外源Eth只是小幅度提高了红丽海棠茎尖冻后存活率,而在Loading环节添加外源SNP冻后存活率反而下降了,SNP通过抑制PCD信号分子NO而其作用,而NO调控PCD具有双重作用,可以促进PCD的发生[29, 30],也可以抑制PCD的发生[31, 32],而添加环节对NO在PCD发生的作用也十分关键,如在春石斛类原球茎中预培养环节导入NO供体(SNP)促进了PCD的发生,降低了存活率,而在Loading环节添加则抑制了PCD的发生[7],故推测外源SNP对不同的植物和不同添加环节可能有不同的影响效果。
4. 结论
红丽海棠茎尖采用以下程序可以实现超低温保存:取组培苗4~5 mm带顶芽茎段,接种在含0.7 mol·L−1蔗糖的预培养基,4 ℃冰箱培养2 d;切取1.5~2.0 mm的茎尖,Loading溶液室温下处理20 min;PVS2溶液0 ℃下处理90 min,可存入液氮保存;需用时将其取出放入38 ℃水浴快速化冻1 min;在室温下Unloading溶液震荡洗涤2次,每次10 min;进行恢复培养,存活率为66.58%,恢复生长率为16.67%。在Loading、PVS2和Unloading环节导入适宜浓度的抗氧化物质CAT、GSH、AsA可明显提高冻后存活率,最佳添加量分别为200 U·ml−1、0.04 μmol·L−1和400 μmol·L−1,较对照存活率分别提高了20.28%、27.61%和6.75%,而添加PCD抑制剂没有显示明显作用
-
![]()
图 1 质粒p-S(A-D)-His构建模式
A:质粒p-S(A-D)-His图谱;B:A位点与D位点连接示意图;C:A位点与D位点连接碱基序列图。
Figure 1. The model diagram of plasmid p-S(A-D)-His construction
A: the plasmid p-S(A-D)-His profile; B: the diagram of the connection between site A and site D; C: the sequence diagram of connecting bases at sites A and D.
![]()
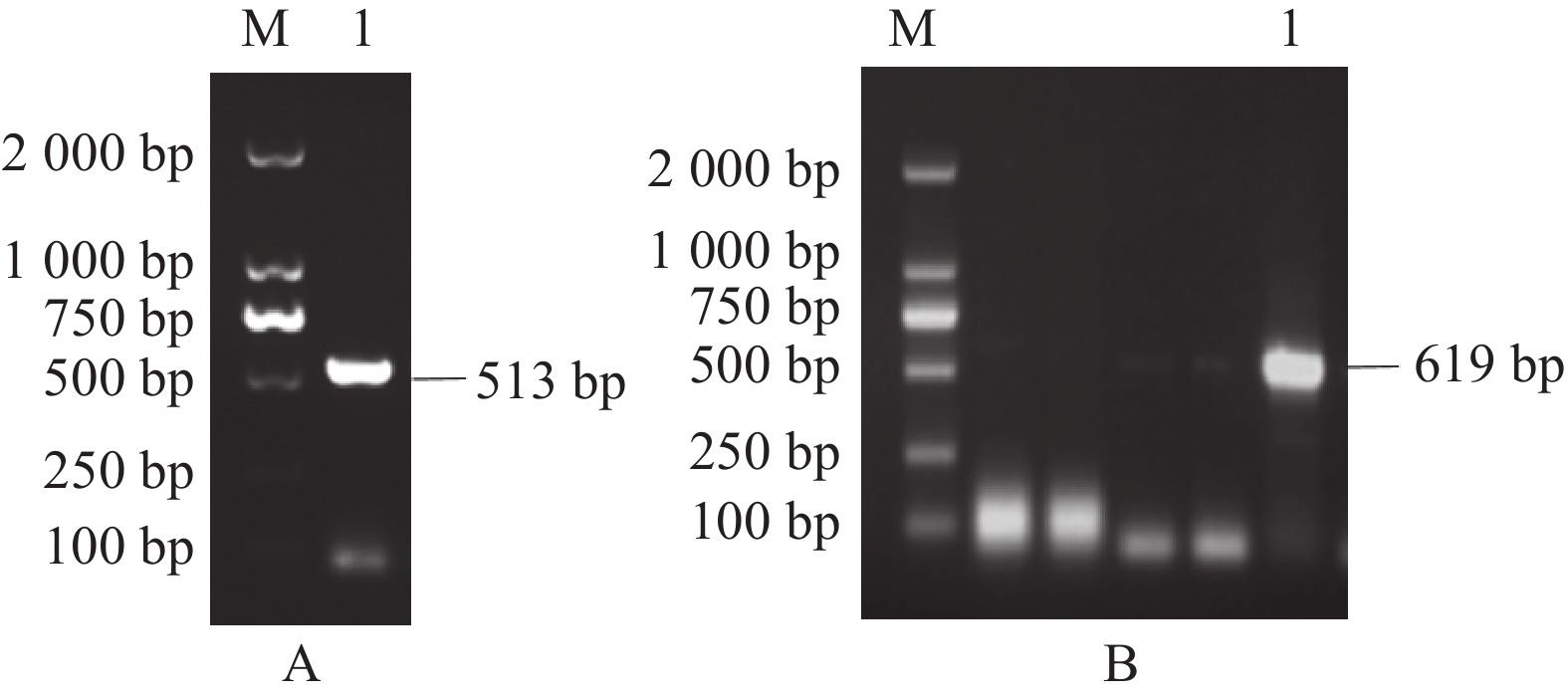
图 2 TGEV S基因A、D位点扩增结果
A:A位点扩增结果;M为DNA分子质量标准DL2000,1为扩增的A位点。B:D位点扩增结果;M为DNA分子质量标准DL2000,1为扩增的D位点。
Figure 2. Amplified A and D sites of TGEV S gene
A: amplification results of A site; M was DNA Marker DL2000, 1 was the PCR-amplified A site. B: amplification results of D site; M was DNA Marker DL2000, 1 was the PCR-amplified D site.
![]()
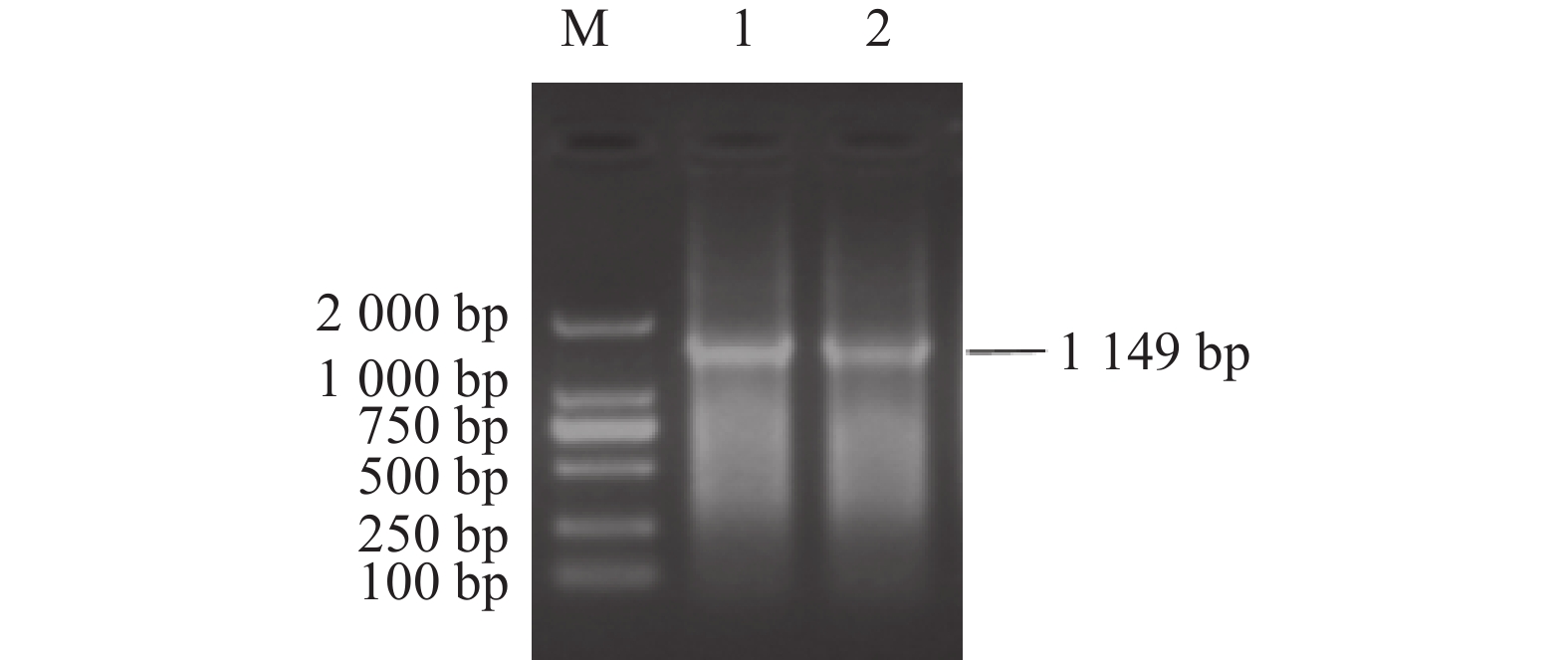
图 3 TGEV N基因扩增结果
M:DNA分子质量标准DL2000;1、2:扩增的N基因。
Figure 3. Amplified N gene of TGEV
M: DNA marker DL2000; 1, 2: PCR-amplified N gene.
![]()
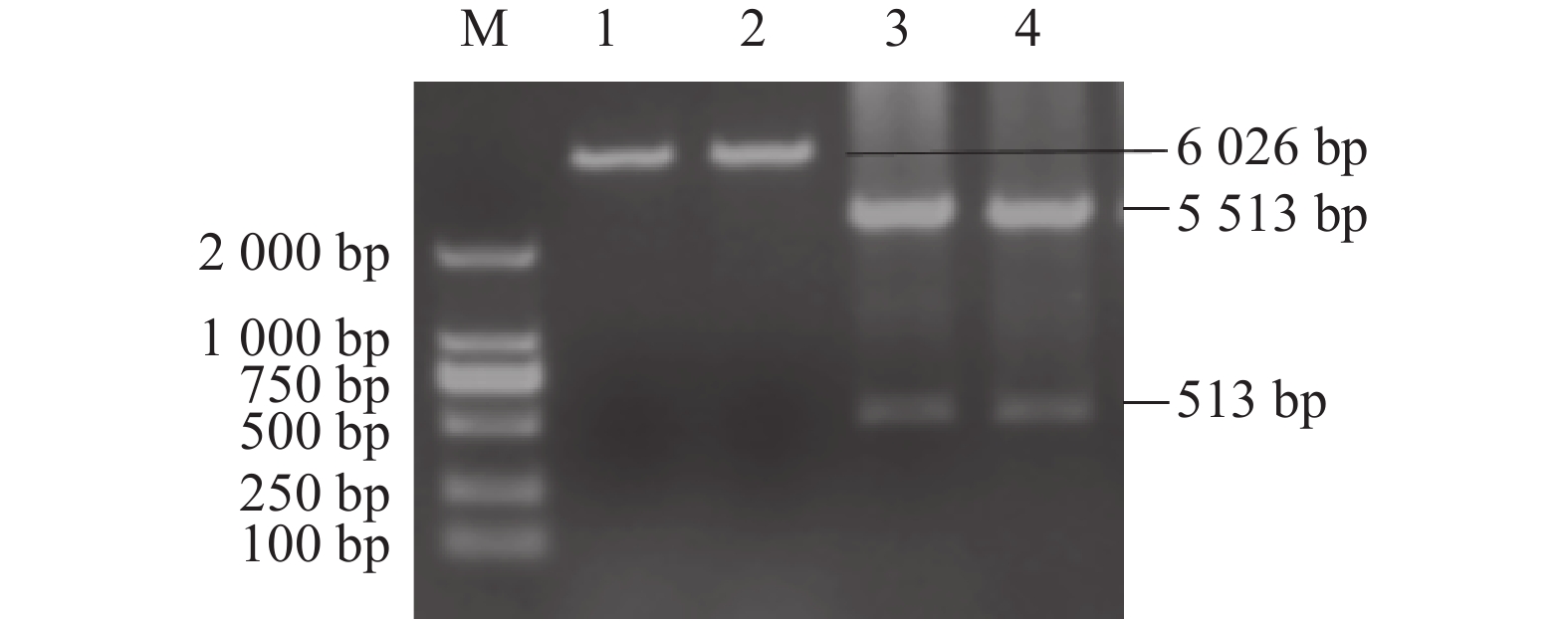
图 4 S基因A位点重组载体p-S(A)-His的构建与鉴定
M:DNA分子质量标准DL2000;1、2:重组载体p-S(A)-His的单酶切;3、4:重组载体p-S(A)-His的双酶切。
Figure 4. Construction and identification of p-S (A)-His and p-S (D)-His at A and D sites of S gene
M: DNA marker DL2000; M2: DNA marker DL10000; 1: single enzyme digestion of recombinant vector p-S (A)-His; 2: single enzyme digestion of recombinant vector p-S (D)-His; 3 and 4: double digestion of recombinant vector p-S (A)-His.
![]()
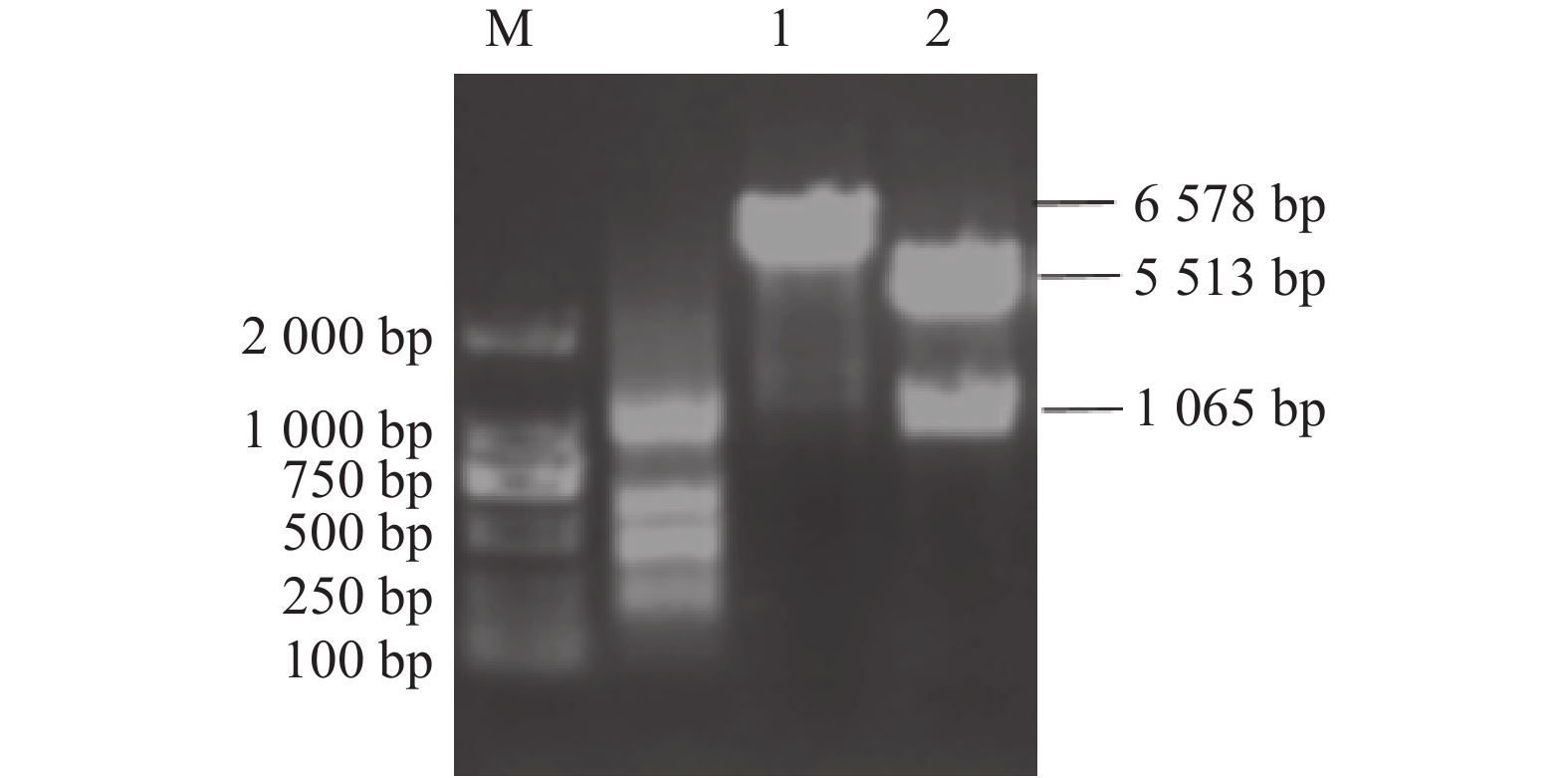
图 5 重组载体p-S(A-D)-His的构建与鉴定
M:DNA分子质量标准DL2000; 1:重组载体p-S(A-D)-His的单酶切;2:重组载体p-S(A-D)-His的双酶切。
Figure 5. Construction and identification of recombinant vectors p-S (A-D)-His
M: DNA marker DL2000; 1: single enzyme digestion of recombinant vector p-S(A-D)-His; 2: double digestion of recombinant vector p-S (A-D)-His.
![]()
图 6 重组载体p-N-His的构建与鉴定
M:DNA分子质量标准DL5000;1:重组载体p-N-His的双酶切。
Figure 6. Construction and identification of recombinant vectors p-N-His
M: DNA marker DL5000; 1: double digestion of recombinant vector p-N-His.
![]()
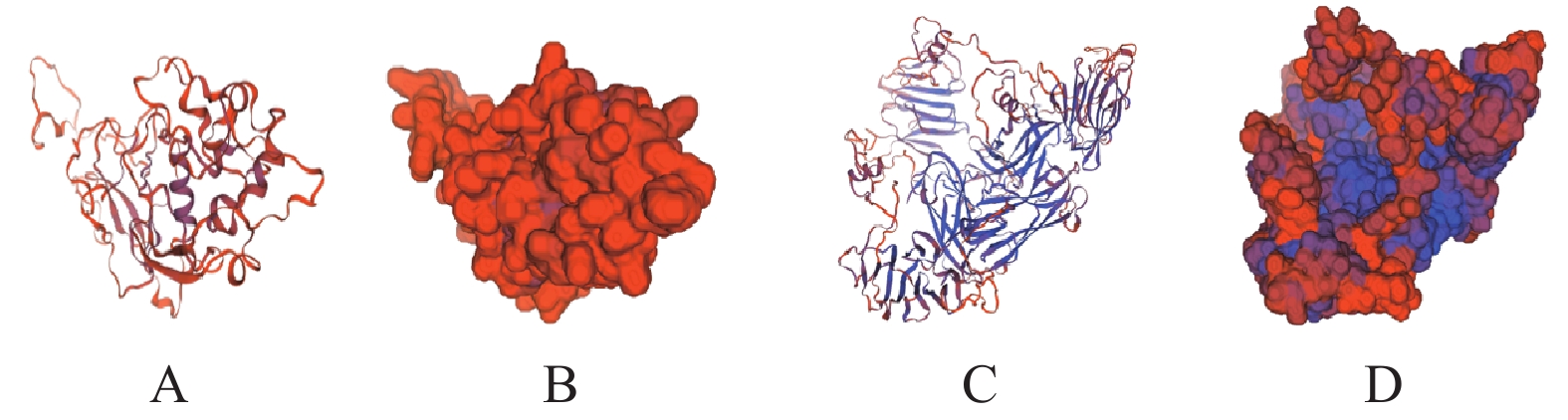
图 7 S(A-D)蛋白、N蛋白三级结构同源建模
A:N蛋白结构图;B:N蛋白表面构象图;C:S(A-D)蛋白结构图;D:S(A-D)蛋白表面构象图。
Figure 7. Homologous modeling on tertiary structures of S (A-D) and N protein
A: Structure of N protein; B: image of N protein structure; C: image of S (A-D) protein structure; D: image of S (A-D) protein structure.
![]()
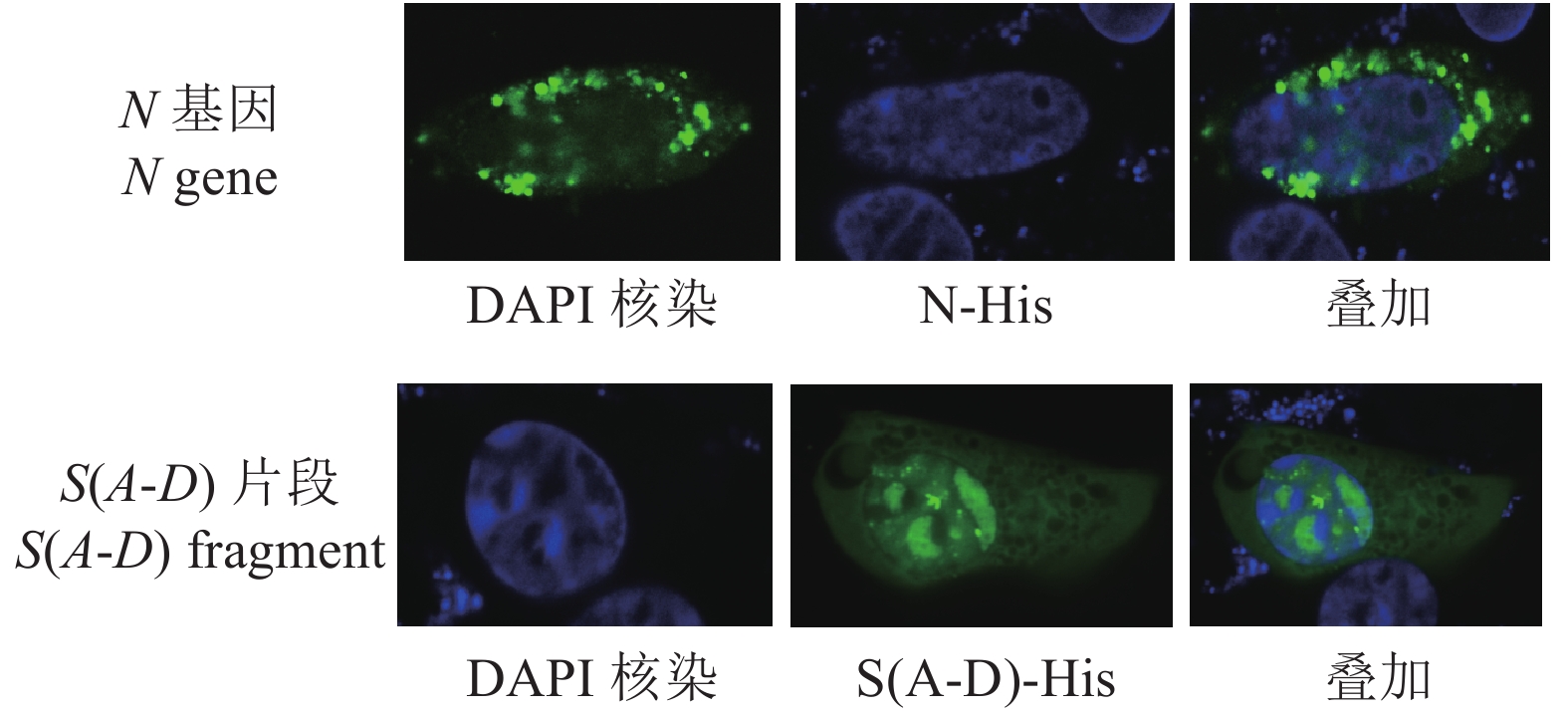
图 8 N基因、S(A-D)片段表达的间接免疫荧光试验
Figure 8. Expressions of N and S (A-D) fragments verified by indirect immunofluorescence assay
表 1 S基因A位点、D位点和N基因扩增引物序列
Table 1 Sequences of primers for amplifications of A and D sites in S gene and N gene
基因
Gene引物
Primers序列(5′−3′)
Sequence(5′−3′)酶切位点
Restriction enzyme cutting siteS基因A位点
A-site in S geneP1 CGCGGATCCATGTTAGTTACCAAACAGCCGT Bam H I P2 CCGGAATTCTATTGTCCAGAAAACGTCAC Eco R I S基因D位点
D-site in S geneP3 CCGGAATTCAAGTTGAAAACACAGCTATT Eco R I P4 TGCTCTAGA ACTATTATCAGACGGTACACC Xba I N基因
N GeneP5 CGCGGATCCATGGCCAACCAGGGAC Bam H I P6 CCGGAATTCGTTCGTTACCTCATCAATT Eco R I 表中加下划线的碱基序列为酶切位点序列。
Cleavage sites are underlined. 下载: 导出CSV
下载: 导出CSV
表 2 小鼠分组及免疫程序
Table 2 Groups and procedures of mice immunization
组别
Group疫苗载体种类
Vaccine carrier type免疫剂量
Immunizing dose/μg小鼠数量
Number of mice免疫时间
Immune frequency免疫位置
Immune site1 p-S(A-D)-His 240 6 第1、7、21天 脚底板 2 p-N-His 240 6 第1、7、21天 脚底板 3 p-S(A-D)-His + p-N-His 120+120 6 第1、7、21天 脚底板 4 pCDNA3.1-His-C 240 6 第1、7、21天 脚底板
下载: 导出CSV
表 3 S(A-D)蛋白和N蛋白亚细胞定位预测
Table 3 Predicted subcellular localization of S (A-D) and N proteins
组别
Group亚细胞定位
Subcellular localization可能性
Possibility/%S(A-D)蛋白
S(A-D) protein细胞质 Cytoplasm 34.8 线粒体 Mitochondria 17.4 细胞核 Cell nucleus 13.0 质膜 Plasmalemma 13.0 内质网 Endoplasmic reticulum 8.7 N蛋白
N protein细胞核 Cell nucleus 65.2 线粒体 Mitochondria 17.4 细胞质 Cytoplasm 13.0 溶酶体 Lysosome 4.3
下载: 导出CSV
表 4 S(A-D)蛋白和N蛋白的B细胞抗原表位预测
Table 4 Predicted B cell epitope of S (A-D) and N proteins
蛋白
Protein序号
Number起始位点
Start site结束位点
End site序列
Amino acid sequenceS(A-D)蛋白
S(A-D)protein1 33 43 FDQCNGAVLNN 2 55 62 TTNVQSGK 3 86 101 DSSFFSYGEIPFGVTD 4 195 211 NLNNGFYPVSSSEVGLV 5 233 250 LGMKRSGYGQPIASTLSN 6 281 295 ALWDNIFKRNCTDVL 7 306 318 CPFSFDKLNNYLT N蛋白
N protein1 4 32 QGQRVSWGDESTKTRGRSNSRGRKSNNIP 2 43 87 QGSKFWNLCPRDFVPNGIGNRDQQIGYWNRQTRYRMVKGQRKELP 3 101 108 ADAKFKDK 4 118 146 DGAMNKPTTLGSRGANNESKALKFDGKVP 5 150 189 QLEVNQSRDNSRSRSQSRSRSRNRSQSRGRQQSNNKKDDS 6 201 244 LGVDTEKQQQRSRSKSKERSNSKTRDTTPKNENKHTWKRTAGKG 7 251 271 GARSSSANFGDSDLVANGSSA 8 316 378 DPKTEQFLQQINAYARPSEVAKEQRKRKSRSKSAERSEQEVVPDALIENYTDVFDDTQVEIID
下载: 导出CSV
表 5 免疫小鼠抗TGEV血清IgG间接ELISA检测
Table 5 Anti-TGEV serum IgG in immunized mice detected by indirect ELISA
组别
Group疫苗载体种类
Vaccine carrier type免疫后不同时间抗TGEV血清IgG水平(OD450 nm)
Anti-TGEV serum IgG levels at different time after immunization (OD450 nm)0天
0 days14天
14 days28天
28 days42天
42 days1 p-S(A-D)-His 0.133±0.011 a 0.218± 0.0075 b0.243± 0.0060 c0.401± 0.0100 c2 p-N-His 0.138±0.016 a 0.244± 0.0025 a0.344± 0.0070 a0.504± 0.0141 a3 p-S(A-D)-His + p-N-His 0.142± 0.0050 a0.225± 0.0021 b0.300± 0.0050 b0.471± 0.0075 b4 pCDNA3.1-His-C 0.134± 0.0089 a0.144± 0.0076 c0.141± 0.0069 c0.139± 0.0020 d同列数据小写字母完全不同表示差异显著(P<0.05),含相同小写字母表示差异不显著(P>0.05)。
Data with different lowercase letters on same column indicate significant differences at P<0.05; those with same lowercase letters indicate no significant differences at P>0.05.
下载: 导出CSV
-
[1] 张羽欣, 王树茂, 段宏勇, 等. 猪传染性胃肠炎病毒TaqMan实时荧光定量PCR检测方法的建立与应用 [J]. 中国兽医科学, 2024, 54(4):479−484. ZHANG Y X, WANG S M, DUAN H Y, et al. Establishment and application of TaqMan real-time quantitative PCR for detection of porcine transmissible gastroenteritis virus [J]. Chinese Veterinary Science, 2024, 54(4): 479−484. (in Chinese)
[2] JI Z Y, DONG H, JIAO R X, et al. The TGEV membrane protein interacts with HSC70 to direct virus internalization through clathrin-mediated endocytosis [J]. Journal of Virology, 2023, 97(4): e0012823. DOI: 10.1128/jvi.00128-23
[3] PU J N, CHEN D W, TIAN G, et al. All-trans retinoic acid attenuates transmissible gastroenteritis virus-induced inflammation in IPEC-J2 cells via suppressing the RLRs/NF-κB signaling pathway [J]. Frontiers in Immunology, 2022, 13: 734171. DOI: 10.3389/fimmu.2022.734171
[4] NIU Z, XU S S, ZHANG Y L, et al. Transmissible gastroenteritis virus nucleocapsid protein interacts with Na+/H+ exchanger 3 to reduce Na+/H+ exchanger activity and promote piglet diarrhea [J]. Journal of Virology, 2022, 96(22): e0147322. DOI: 10.1128/jvi.01473-22
[5] QIAN J T, LI M J, FENG Y F, et al. Genetic epidemiology of porcine transmissible gastroenteritis virus based on whole genome and S gene sequences[C]//2021 IEEE 9th International Conference on Bioinformatics and Computational Biology (ICBCB). May 25-27, 2021, Taiyuan, China. IEEE, 2021: 148-151.
[6] 郝振业. 猪传染性胃肠炎病毒反向遗传操作系统的构建及附属蛋白3(ORF3)的定位及功能研究[D]. 哈尔滨: 东北农业大学, 2023. HAO Z Y. Construction of reverse genetic operating system of porcine transmissible gastroenteritis virus and study on location and function of accessory protein 3(ORF3)[D]. Harbin: Northeast Agricultural University, 2023. (in Chinese)
[7] 王艳春. 猪传染性胃肠炎病毒S基因A位点杆状病毒表达及初步应用[D]. 扬州: 扬州大学, 2020. WANG Y C. Expression of baculovirus at site A of S gene of porcine infectious gastroenteritis virus and its preliminary application[D]. Yangzhou: Yangzhou University, 2020. (in Chinese)
[8] 张海燕. PEDV和TGEV的S蛋白融合抗原表位核酸疫苗的研究[D]. 武汉: 华中农业大学, 2019. ZHANG H Y. Study on S protein fusion epitope nucleic acid vaccine of PEDV and TGEV[D]. Wuhan: Huazhong Agricultural University, 2019. (in Chinese)
[9] LI X L, LI P C, CAO L Y, et al. Porcine IL-12 plasmid as an adjuvant improves the cellular and humoral immune responses of DNA vaccine targeting transmissible gastroenteritis virus spike gene in a mouse model [J]. Journal of Veterinary Medical Science, 2019, 81(10): 1438−1444. DOI: 10.1292/jvms.18-0682
[10] 师一鸣. TGEV单克隆抗体的制备及检测方法的建立[D]. 哈尔滨: 东北农业大学, 2019. SHI Y M. Preparation of TGEV monoclonal antibody and establishment of detection method[D]. Harbin: Northeast Agricultural University, 2019. (in Chinese)
[11] 李雅静, 宫强. 铜绿假单胞菌oprH基因DNA疫苗的构建与检测 [J]. 现代畜牧兽医, 2022(11):19−23. DOI: 10.3969/j.issn.1672-9692.2022.11.lnxmsy202211006 LI Y J, GONG Q. Construction and detection of Pseudomonas aeruginosa oprH gene DNA vaccine [J]. Modern Journal of Animal Husbandry and Veterinary Medicine, 2022(11): 19−23. (in Chinese) DOI: 10.3969/j.issn.1672-9692.2022.11.lnxmsy202211006
[12] 杨鹏, 吴燕, 岳筠, 等. 绵羊肺炎支原体P113蛋白C末端基因真核表达载体的构建及其小鼠免疫应答 [J]. 中国兽医学报, 2022, 42(3):496−501, 521. YANG P, WU Y, YUE J, et al. Construction of eukaryotic expression vector for C terminal gene of Mycoplasma ovipneumoniae P113 protein and its immune response in mice [J]. Chinese Journal of Veterinary Science, 2022, 42(3): 496−501, 521. (in Chinese)
[13] 姚思, 杨洁琼, 杨雨欣, 等. 结核分枝杆菌ESAT6-Fc DNA疫苗的免疫效应评价 [J]. 中国生物制品学杂志, 2023, 36(8):897−901. YAO S, YANG J Q, YANG Y X, et al. Evaluation of immune effect of ESAT6-Fc DNA vaccine against Mycobacterium tuberculosis [J]. Chinese Journal of Biologicals, 2023, 36(8): 897−901. (in Chinese)
[14] 黄小波, 杨恒, 曹三杰, 等. 猪传染性胃肠炎病毒S-N融合双基因疫苗的构建及其免疫原性分析 [J]. 中国兽医科学, 2012, 42(8):848−853. HUANG X B, YANG H, CAO S J, et al. Construction and immunogenicity analysis of the S-N fusion gene vaccine against porcine transmissible gastroenteritis virus [J]. Chinese Veterinary Science, 2012, 42(8): 848−853. (in Chinese)
[15] WANG G, LIANG R, LIU Z W, et al. The N-terminal domain of spike protein is not the enteric tropism determinant for transmissible gastroenteritis virus in piglets [J]. Viruses, 2019, 11(4): 313. DOI: 10.3390/v11040313
[16] 韩郁茹, 石达, 张记宇, 等. 猪急性腹泻综合征冠状病毒RT-LAMP快速检测方法的建立与应用 [J]. 中国预防兽医学报, 2021, 43(1):35−39. HAN Y R, SHI D, ZHANG J Y, et al. Development and application of RT-LAMP method for rapid detection of SADS-CoV [J]. Chinese Journal of Preventive Veterinary Medicine, 2021, 43(1): 35−39. (in Chinese)
[17] 樊杰. 猪传染性胃肠炎病毒N蛋白纳米抗体的制备和基于纳米抗体竞争ELISA的建立[D]. 杨凌: 西北农林科技大学, 2021. FAN J. Preparation of nano-antibody against N protein of transmissible gastroenteritis virus and establishment of competitive ELISA based on nano-antibody[D]. Yangling: Northwest A & F University, 2021. (in Chinese)
[18] ZHANG Y D, ZHANG X H, LIAO X D, et al. Construction of a bivalent DNA vaccine co-expressing S genes of transmissible gastroenteritis virus and porcine epidemic diarrhea virus delivered by attenuated Salmonella typhimurium [J]. Virus Genes, 2016, 52(3): 354−364. DOI: 10.1007/s11262-016-1316-z
[19] 何雷, 董玲娟, 张彦明, 等. 猪传染性胃肠炎病毒ORF7蛋白在ST细胞中定位及其对病毒复制影响的研究 [J]. 中国预防兽医学报, 2020, 42(6):543−548. HE L, DONG L J, ZHANG Y M, et al. The subcellular location of transmissible gastroenteritis virus protein ORF7 and its effect on viral replication [J]. Chinese Journal of Preventive Veterinary Medicine, 2020, 42(6): 543−548. (in Chinese)
[20] 何雷, 贾艳艳, 郁川, 等. 稳定表达猪传染性胃肠炎病毒N蛋白的ST细胞株的构建及其亚细胞定位 [J]. 中国预防兽医学报, 2016, 38(2):101−104. DOI: 10.3969/j.issn.1008-0589.2016.02.04 HE L, JIA Y Y, YU C, et al. Establishment of stably-expressed transmissible gastroenteritis virus N protein cell line and its subcellular location [J]. Chinese Journal of Preventive Veterinary Medicine, 2016, 38(2): 101−104. (in Chinese) DOI: 10.3969/j.issn.1008-0589.2016.02.04
[21] 韩涛涛, 黎露, 唐青海, 等. 不同佐剂对猪传染性胃肠炎病毒S蛋白和猪流行性腹泻病毒S蛋白免疫原性的影响 [J]. 中国农学通报, 2020, 36(30):143−150. DOI: 10.11924/j.issn.1000-6850.casb20191000740 HAN T T, LI L, TANG Q H, et al. Different adjuvants: Effects on S protein immunogenicity of porcine transmissible gastroenteritis virus and porcine epidemic diarrhea virus [J]. Chinese Agricultural Science Bulletin, 2020, 36(30): 143−150. (in Chinese) DOI: 10.11924/j.issn.1000-6850.casb20191000740
[22] 伊立超. PEDV和TGEV受体结合区基因在昆虫杆状病毒系统的表达与免疫原性分析[D]. 延吉: 延边大学, 2022. YI L C. Expression and immunogenicity analysis of PEDV and TGEV receptor binding region genes in insect baculovirus system[D]. Yanji: Yanbian University, 2022. (in Chinese)
-
期刊类型引用(1)
1. 戴惠新,吴金洋,张梦樵,张湘菊,吴丽娜. 喷涂机器人用改性聚天门冬氨酸酯腻子制备及性能测试. 粘接. 2024(12): 42-45 .  百度学术
百度学术
其他类型引用(0)











计量
- 文章访问数: 105
- HTML全文浏览量: 49
- PDF下载量: 34
- 被引次数: 1
